Structural and Computational Biology Unit, European Molecular Biology Laboratory, 69117 Heidelberg, Germany.
Department of Molecular Biology, Max Planck Institute for Biophysical Chemistry, 37077 Gottingen, Germany.
Proc Natl Acad Sci U S A. 2019 May 14;116(20):10048-10057. doi: 10.1073/pnas.1900931116. Epub 2019 Apr 29.
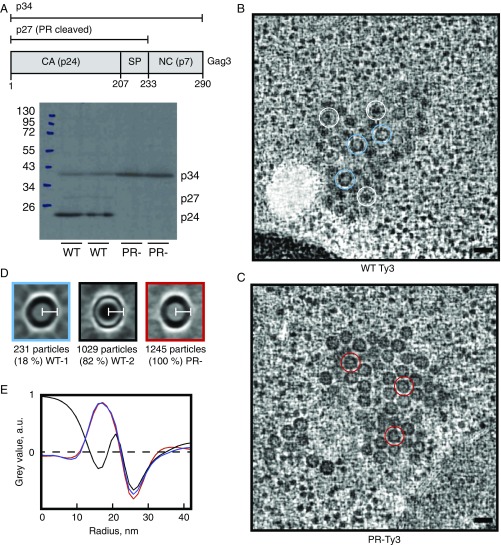
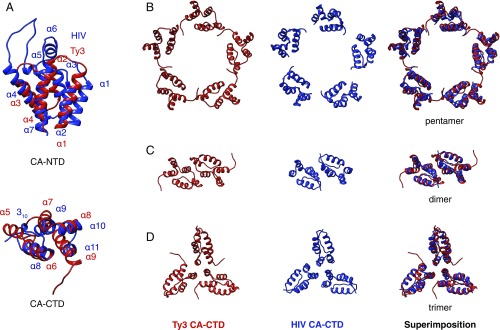
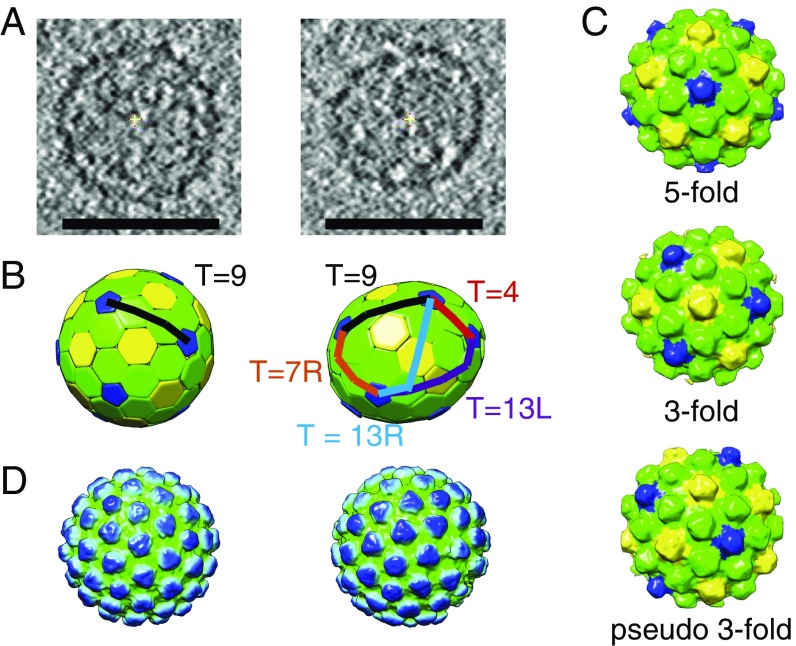
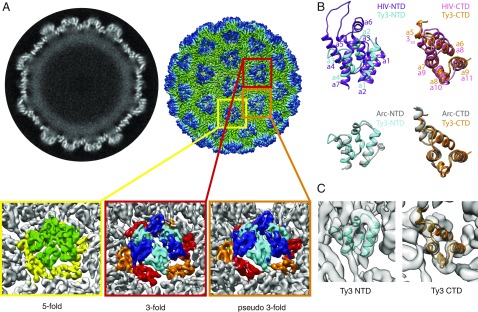
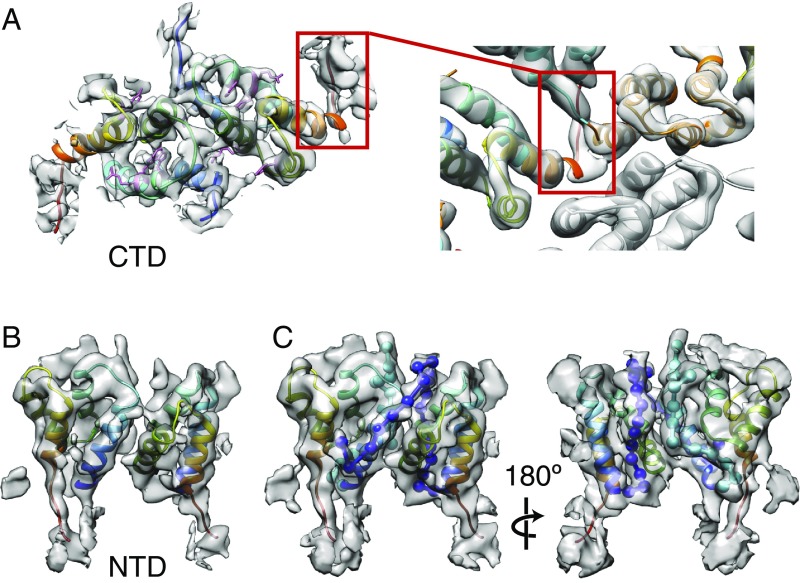
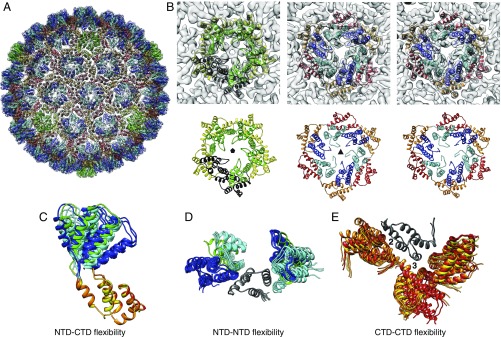
Retroviruses evolved from long terminal repeat (LTR) retrotransposons by acquisition of envelope functions, and subsequently reinvaded host genomes. Together, endogenous retroviruses and LTR retrotransposons represent major components of animal, plant, and fungal genomes. Sequences from these elements have been exapted to perform essential host functions, including placental development, synaptic communication, and transcriptional regulation. They encode a Gag polypeptide, the capsid domains of which can oligomerize to form a virus-like particle. The structures of retroviral capsids have been extensively described. They assemble an immature viral particle through oligomerization of full-length Gag. Proteolytic cleavage of Gag results in a mature, infectious particle. In contrast, the absence of structural data on LTR retrotransposon capsids hinders our understanding of their function and evolutionary relationships. Here, we report the capsid morphology and structure of the archetypal Gypsy retrotransposon Ty3. We performed electron tomography (ET) of immature and mature Ty3 particles within cells. We found that, in contrast to retroviruses, these do not change size or shape upon maturation. Cryo-ET and cryo-electron microscopy of purified, immature Ty3 particles revealed an irregular fullerene geometry previously described for mature retrovirus core particles and a tertiary and quaternary arrangement of the capsid (CA) C-terminal domain within the assembled capsid that is conserved with mature HIV-1. These findings provide a structural basis for studying retrotransposon capsids, including those domesticated in higher organisms. They suggest that assembly via a structurally distinct immature capsid is a later retroviral adaptation, while the structure of mature assembled capsids is conserved between LTR retrotransposons and retroviruses.
逆转录病毒通过获得包膜功能从长末端重复序列 (LTR) 逆转录转座子进化而来,随后重新入侵宿主基因组。内源性逆转录病毒和 LTR 逆转录转座子共同构成了动物、植物和真菌基因组的主要组成部分。这些元件的序列被改编为执行重要的宿主功能,包括胎盘发育、突触通讯和转录调控。它们编码 Gag 多肽,其衣壳域可以寡聚形成病毒样颗粒。逆转录病毒衣壳的结构已得到广泛描述。它们通过全长 Gag 的寡聚化组装不成熟的病毒颗粒。Gag 的蛋白水解切割导致成熟的、感染性的颗粒。相比之下,缺乏关于 LTR 逆转录转座子衣壳的结构数据阻碍了我们对其功能和进化关系的理解。在这里,我们报告了原型 Gypsy 逆转录转座子 Ty3 的衣壳形态和结构。我们在细胞内对不成熟和成熟的 Ty3 颗粒进行了电子断层扫描 (ET)。我们发现,与逆转录病毒不同,这些颗粒在成熟时不会改变大小或形状。对纯化的不成熟 Ty3 颗粒进行冷冻电子断层扫描和冷冻电子显微镜检查,揭示了以前在成熟逆转录病毒核心颗粒中描述过的不规则富勒烯几何形状,以及组装衣壳内衣壳 (CA) C 末端结构域的三级和四级排列,这与成熟的 HIV-1 保守一致。这些发现为研究逆转录转座子衣壳提供了结构基础,包括那些在高等生物中驯化的衣壳。它们表明,通过结构上不同的不成熟衣壳进行组装是逆转录病毒的一个后期适应,而成熟组装衣壳的结构在 LTR 逆转录转座子和逆转录病毒之间是保守的。