Plowman Rebecca, Singh Namit, Tromer Eelco C, Payan Angel, Duro Eris, Spanos Christos, Rappsilber Juri, Snel Berend, Kops Geert J P L, Corbett Kevin D, Marston Adele L
Wellcome Centre for Cell Biology, School of Biological Sciences, University of Edinburgh, Max Born Crescent, Edinburgh, EH9 3BF, UK.
Ludwig Institute for Cancer Research, San Diego Branch, La Jolla, CA, 92093, USA.
Chromosoma. 2019 Sep;128(3):331-354. doi: 10.1007/s00412-019-00700-0. Epub 2019 Apr 30.
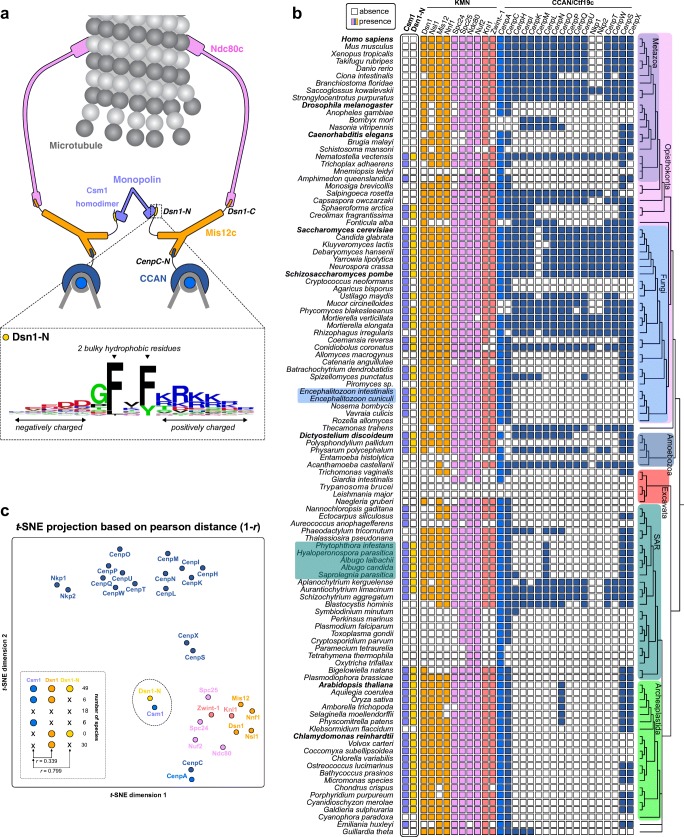
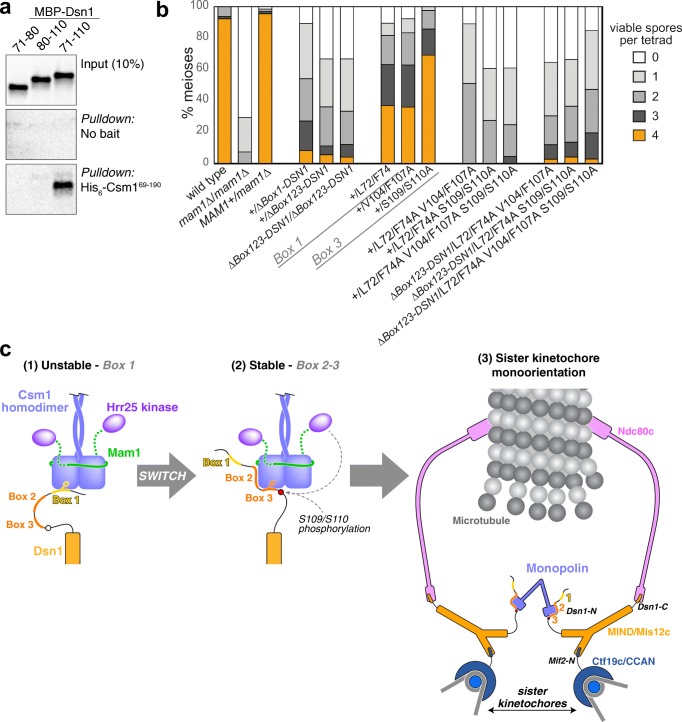
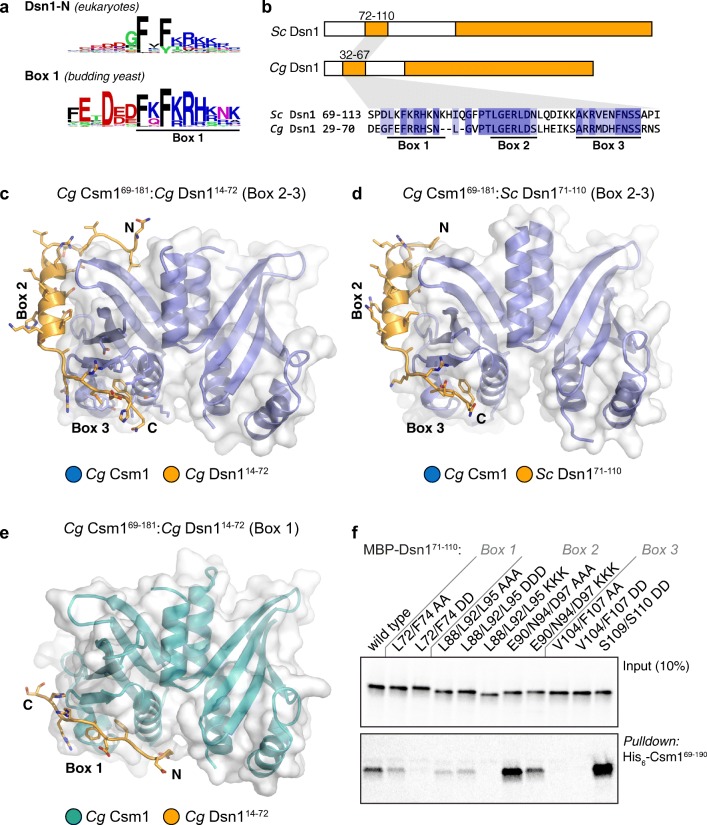
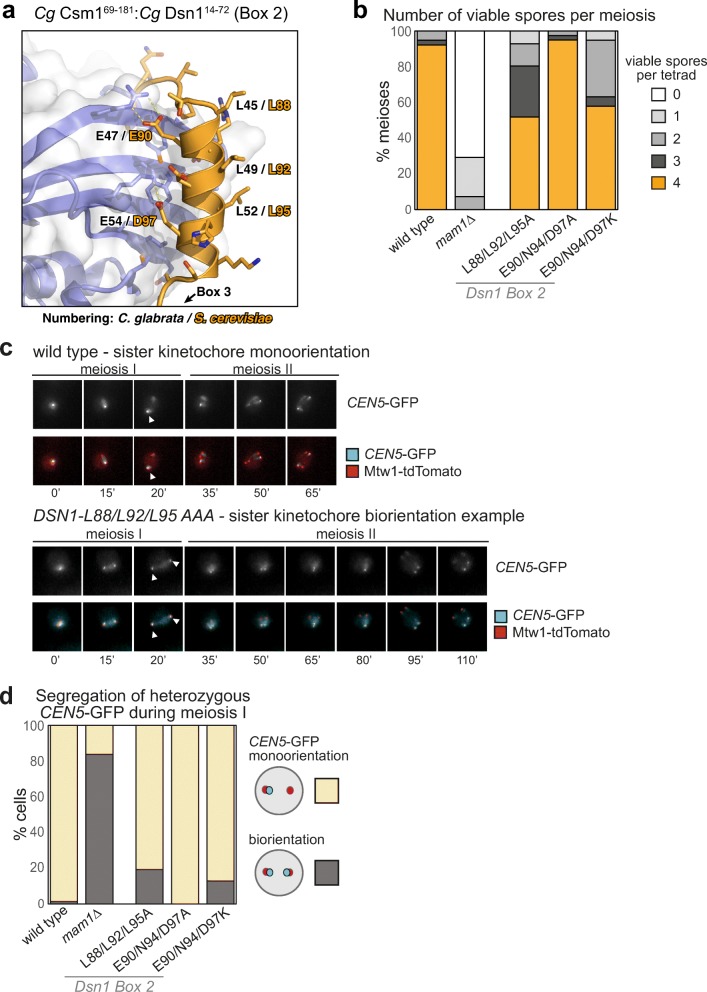
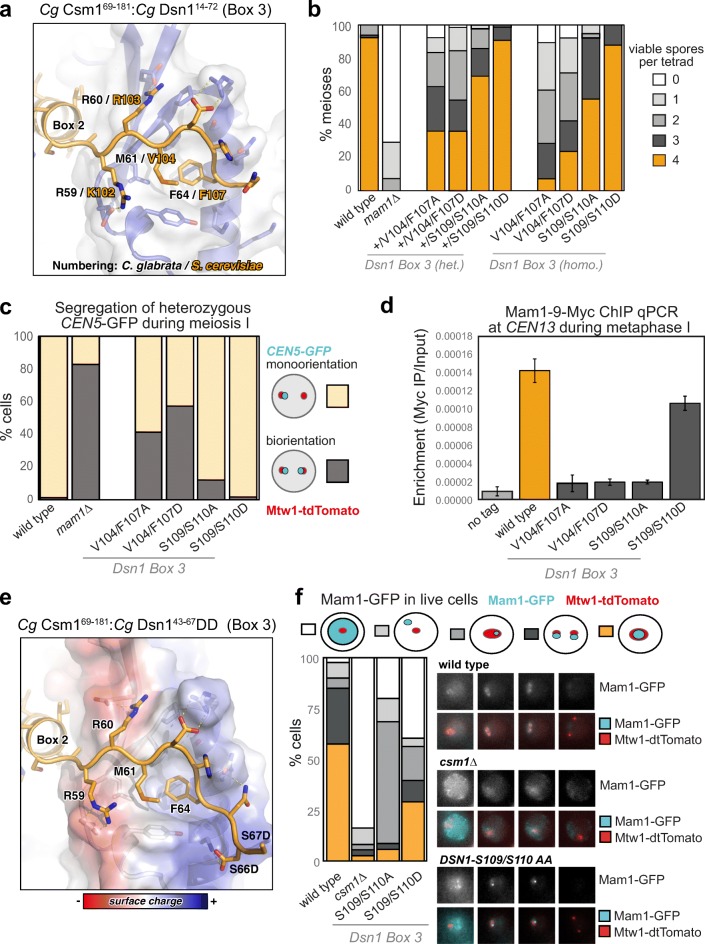
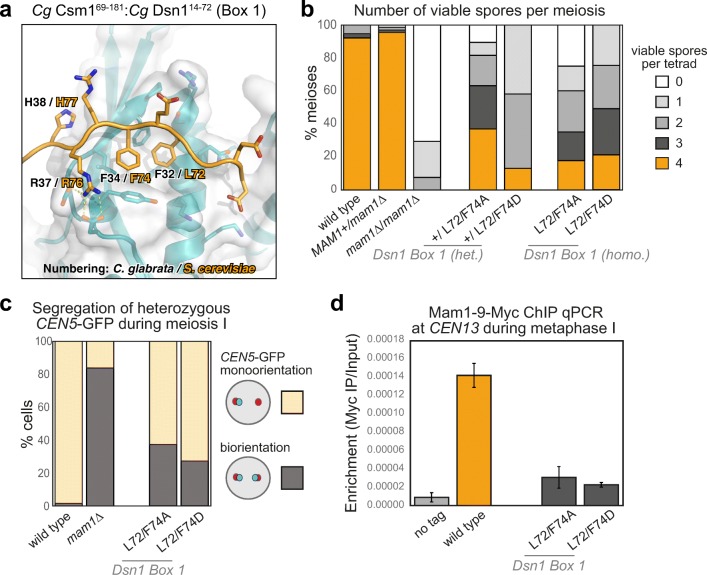
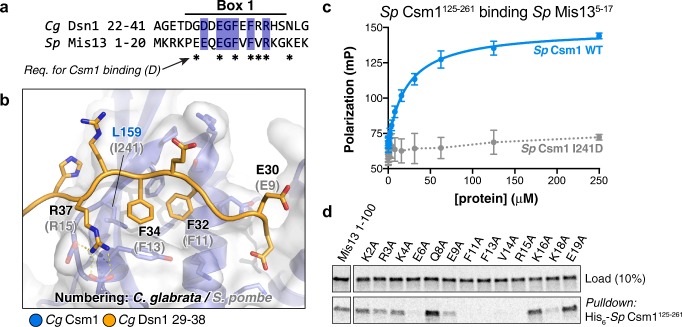
The monopolin complex is a multifunctional molecular crosslinker, which in S. pombe binds and organises mitotic kinetochores to prevent aberrant kinetochore-microtubule interactions. In the budding yeast S. cerevisiae, whose kinetochores bind a single microtubule, the monopolin complex crosslinks and mono-orients sister kinetochores in meiosis I, enabling the biorientation and segregation of homologs. Here, we show that both the monopolin complex subunit Csm1 and its binding site on the kinetochore protein Dsn1 are broadly distributed throughout eukaryotes, suggesting a conserved role in kinetochore organisation and function. We find that budding yeast Csm1 binds two conserved motifs in Dsn1, one (termed Box 1) representing the ancestral, widely conserved monopolin binding motif and a second (termed Box 2-3) with a likely role in enforcing specificity of sister kinetochore crosslinking. We find that Box 1 and Box 2-3 bind the same conserved hydrophobic cavity on Csm1, suggesting competition or handoff between these motifs. Using structure-based mutants, we also find that both Box 1 and Box 2-3 are critical for monopolin function in meiosis. We identify two conserved serine residues in Box 2-3 that are phosphorylated in meiosis and whose mutation to aspartate stabilises Csm1-Dsn1 binding, suggesting that regulated phosphorylation of these residues may play a role in sister kinetochore crosslinking specificity. Overall, our results reveal the monopolin complex as a broadly conserved kinetochore organiser in eukaryotes, which budding yeast have co-opted to mediate sister kinetochore crosslinking through the addition of a second, regulatable monopolin binding interface.
单极蛋白复合体是一种多功能分子交联剂,在粟酒裂殖酵母中,它结合并组织有丝分裂动粒,以防止异常的动粒 - 微管相互作用。在芽殖酵母酿酒酵母中,其动粒结合单个微管,单极蛋白复合体在减数分裂I中交联并单定向姐妹动粒,使同源染色体能够双定向和分离。在这里,我们表明单极蛋白复合体亚基Csm1及其在动粒蛋白Dsn1上的结合位点在整个真核生物中广泛分布,这表明在动粒组织和功能中具有保守作用。我们发现芽殖酵母Csm1结合Dsn1中的两个保守基序,一个(称为Box 1)代表祖先广泛保守的单极蛋白结合基序,另一个(称为Box 2 - 3)可能在增强姐妹动粒交联的特异性方面发挥作用。我们发现Box 1和Box 2 - 3结合Csm1上相同的保守疏水腔,表明这些基序之间存在竞争或交接。使用基于结构的突变体,我们还发现Box 1和Box 2 - 3对于减数分裂中的单极蛋白功能至关重要。我们在Box 2 - 3中鉴定出两个保守的丝氨酸残基,它们在减数分裂中被磷酸化,并且将其突变为天冬氨酸可稳定Csm1 - Dsn1结合,这表明这些残基的磷酸化调控可能在姐妹动粒交联特异性中起作用。总体而言,我们的结果揭示了单极蛋白复合体是真核生物中广泛保守的动粒组织者,芽殖酵母通过添加第二个可调节的单极蛋白结合界面来利用它介导姐妹动粒交联。