Laboratory of Cellular and Molecular Biology,Center for Cancer Research, National Cancer Institute, National Institutes of Health, Bethesda, United States.
Program of Mathematical and Life Sciences, Graduate School of Integrated Science for Life, Hiroshima University, Higashi-Hiroshima, Japan.
Elife. 2019 Jul 10;8:e44752. doi: 10.7554/eLife.44752.
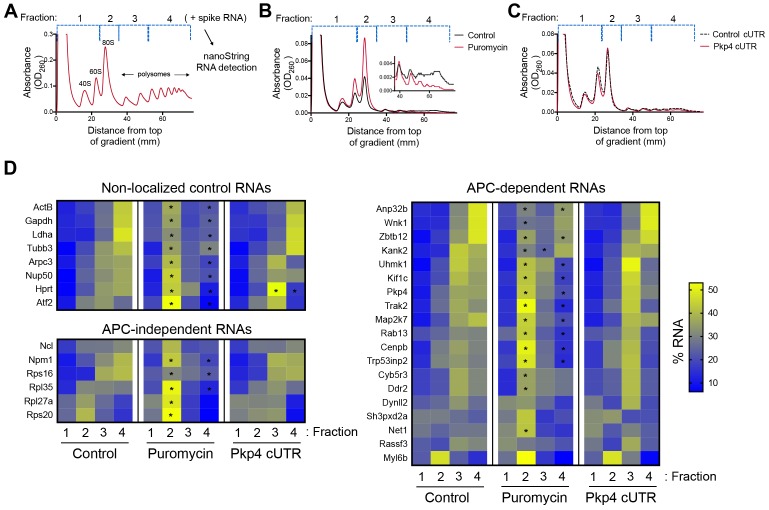
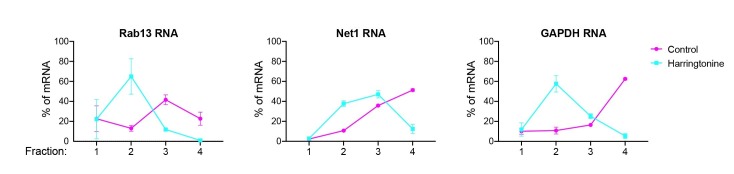
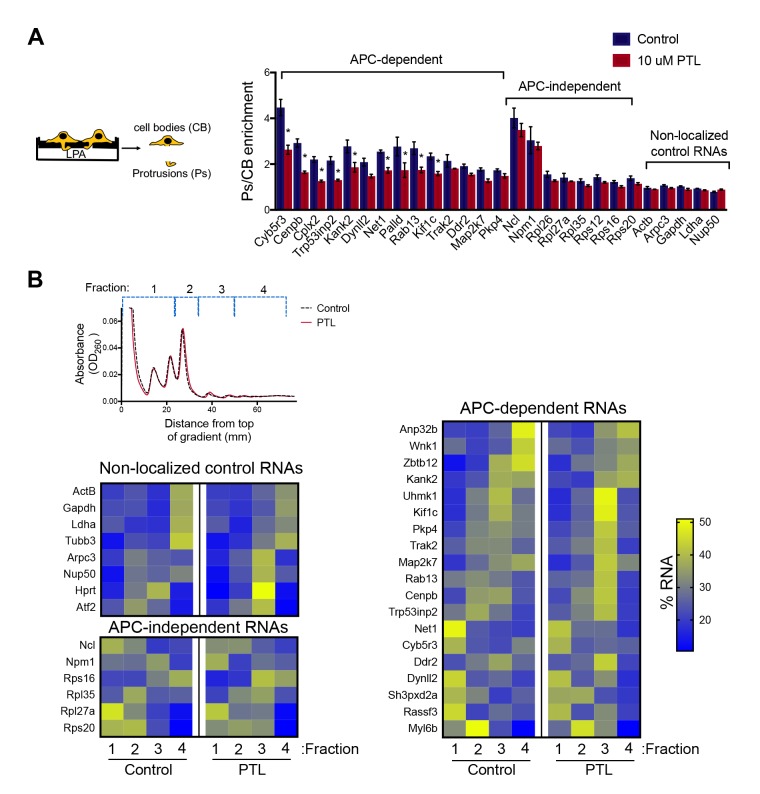
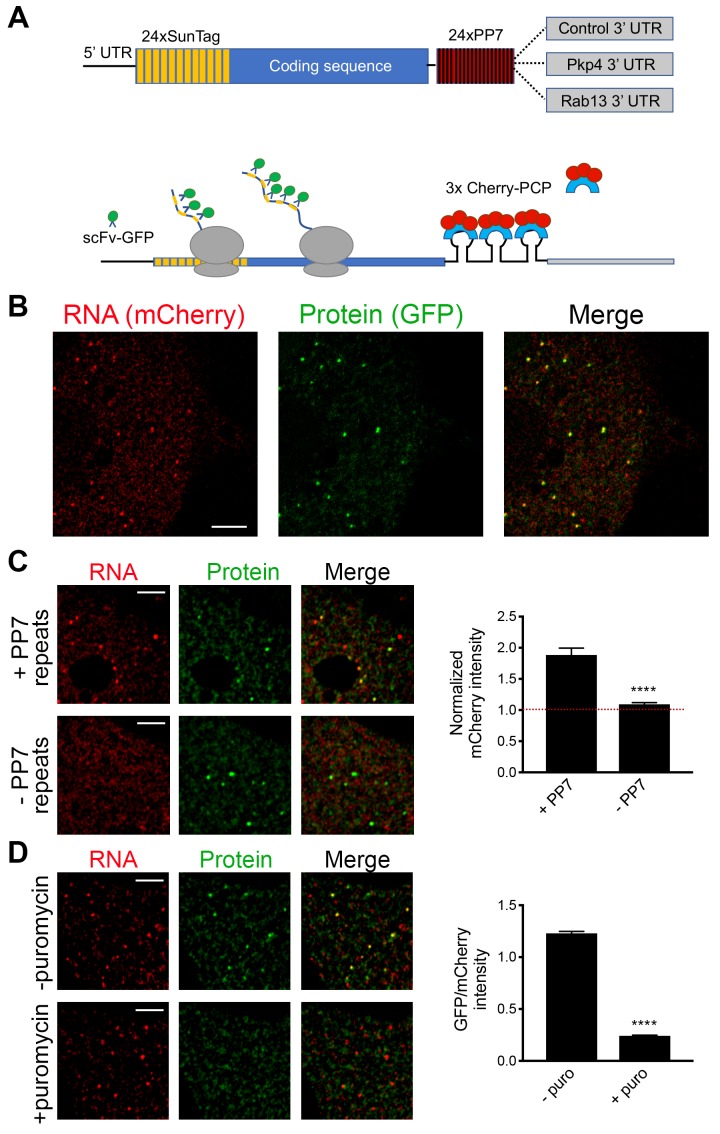
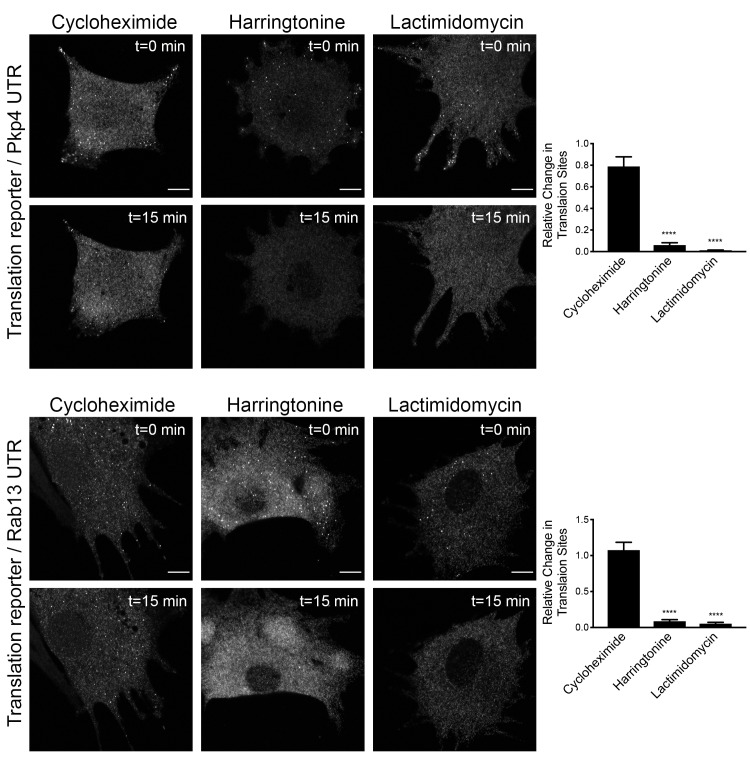
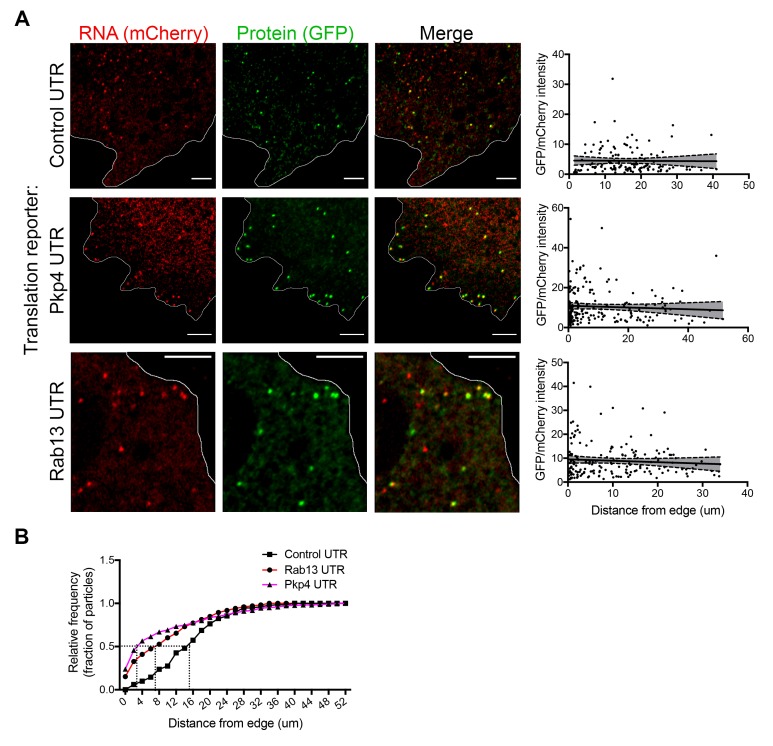
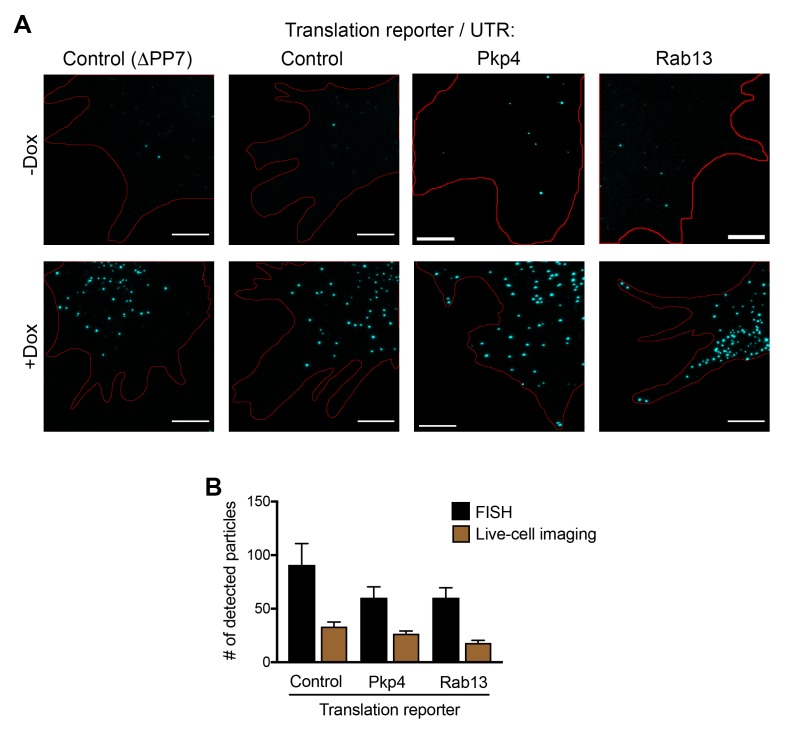
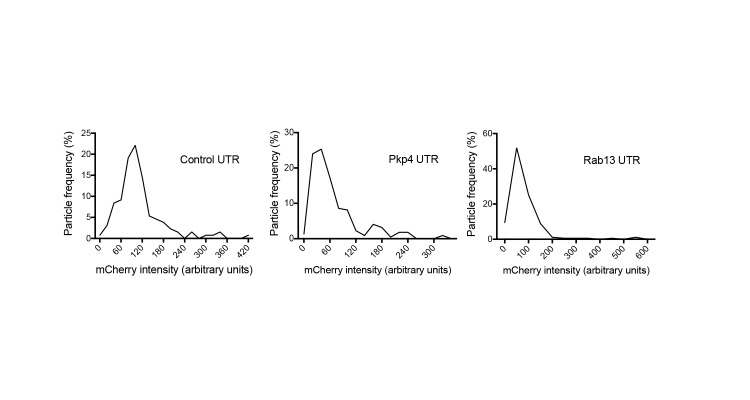
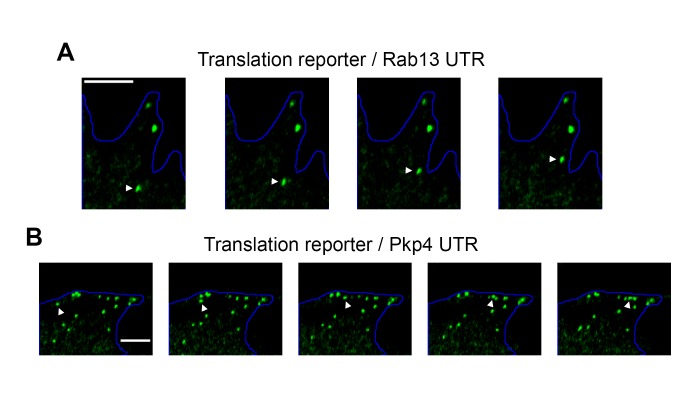
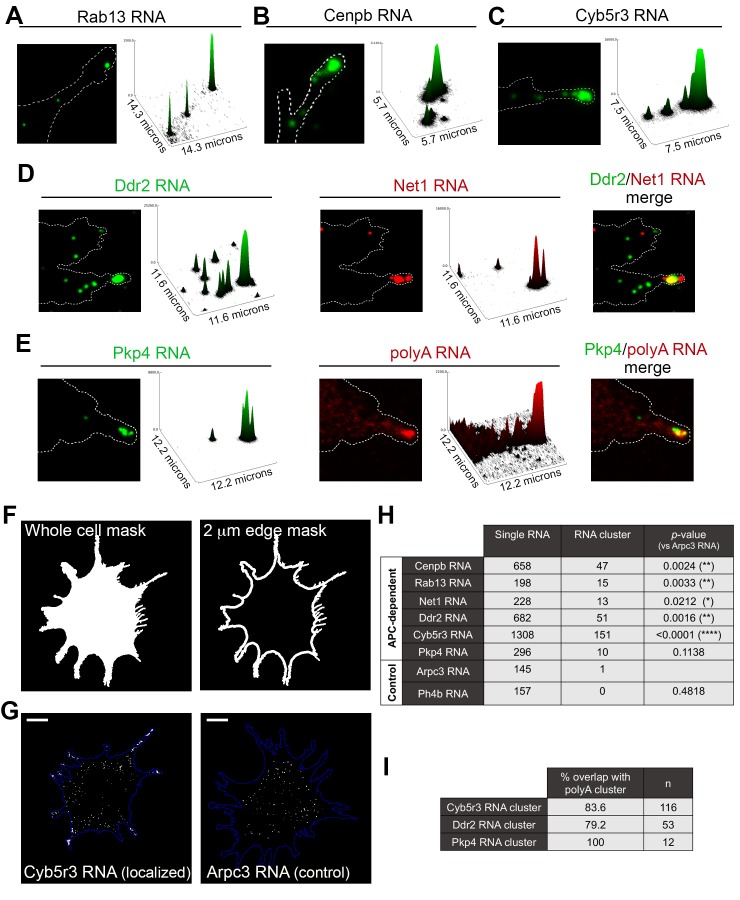
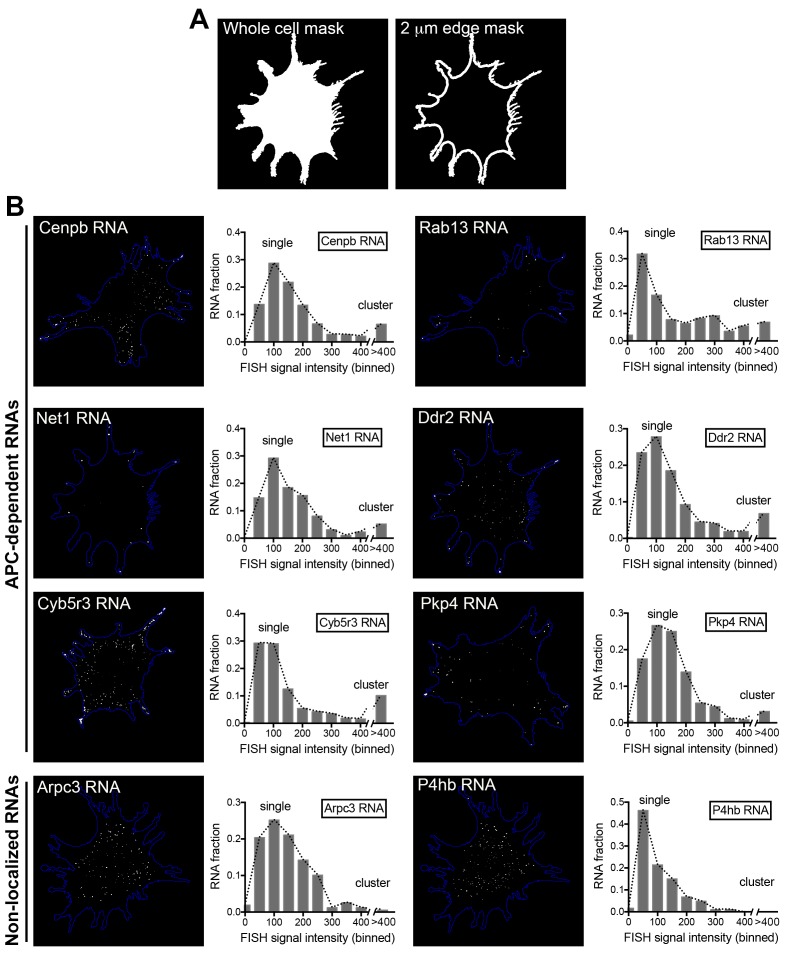
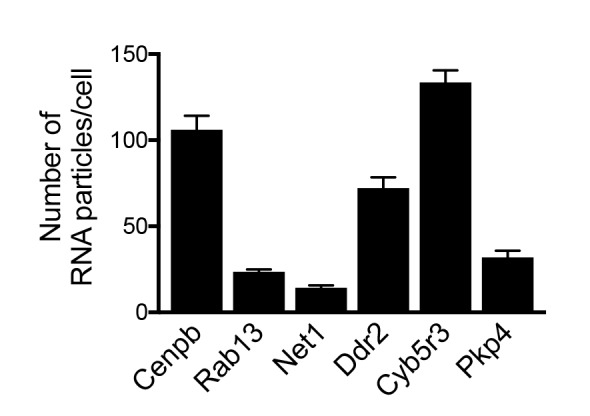
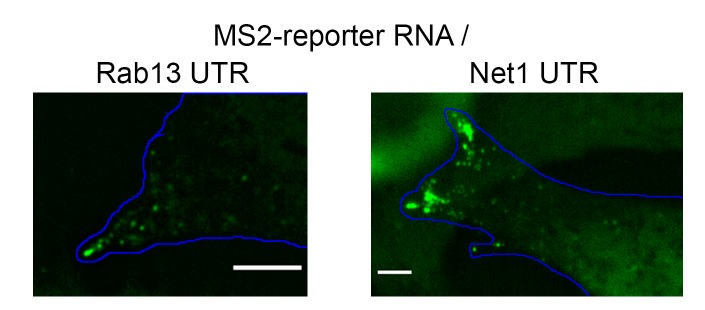
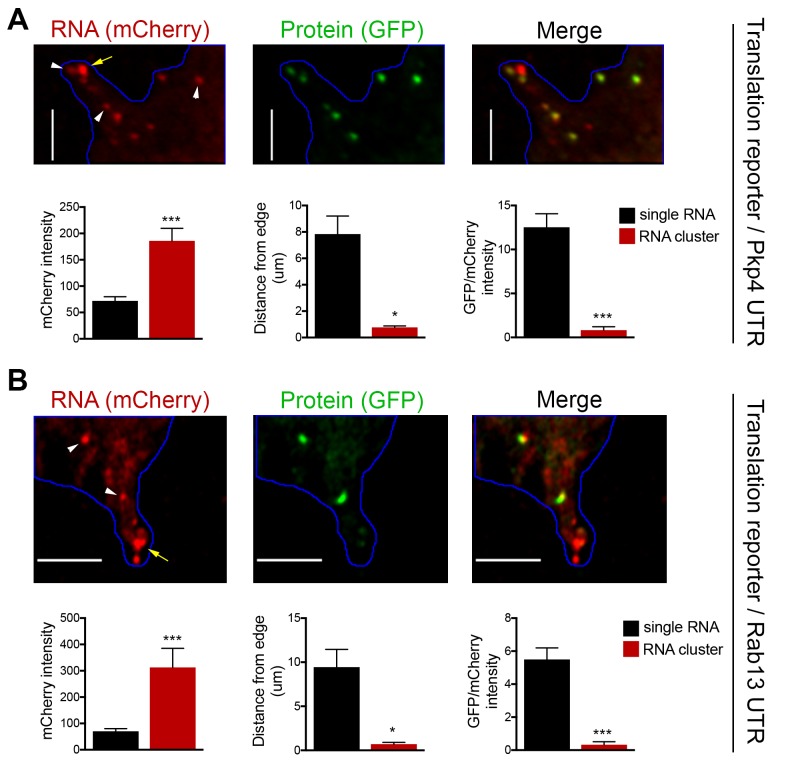
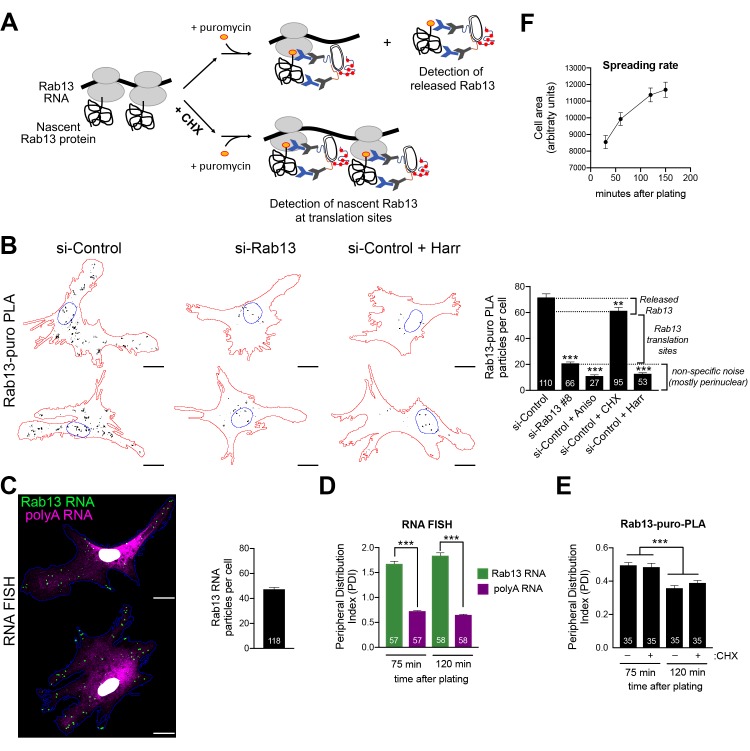
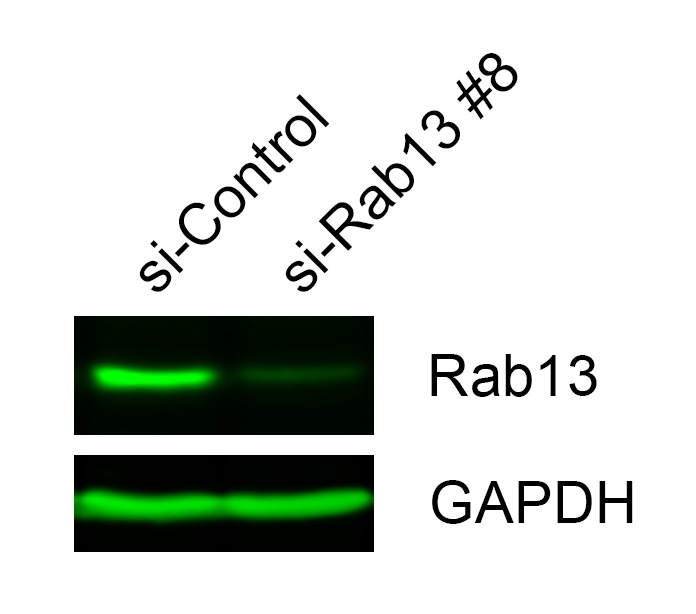
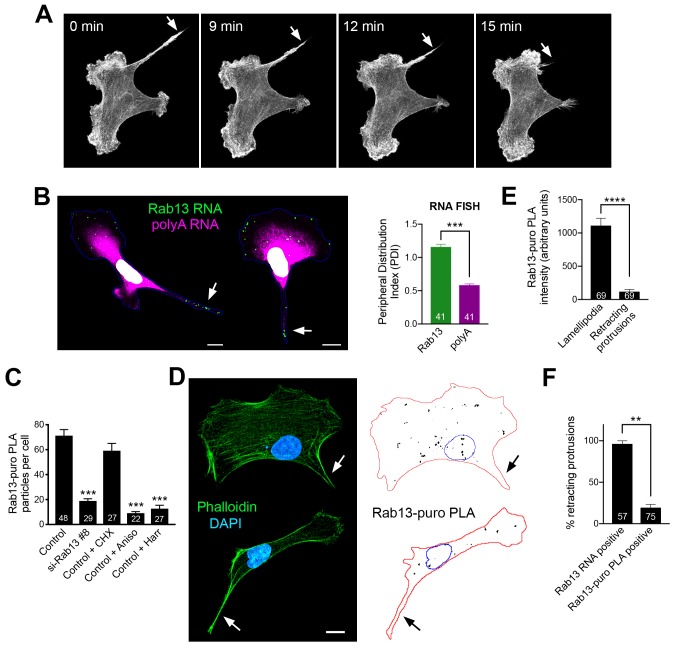
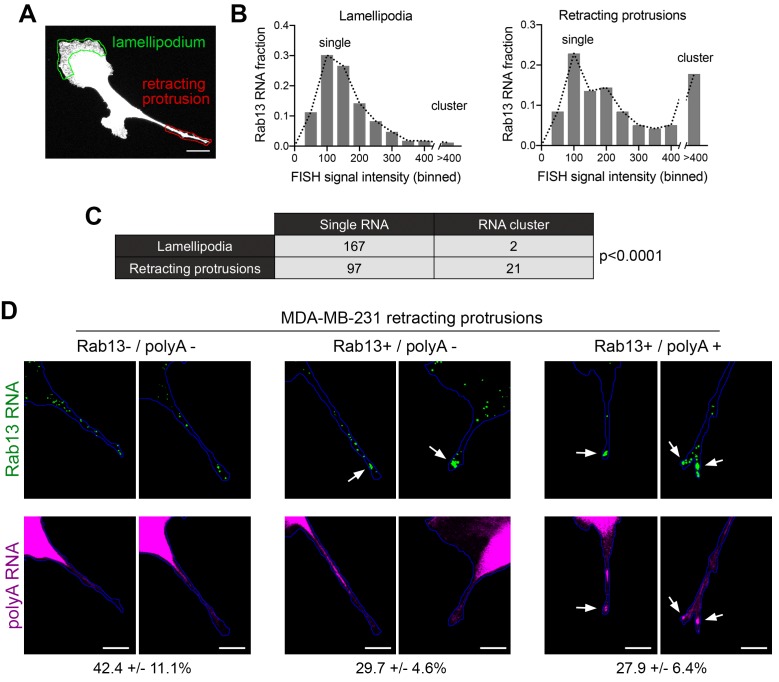
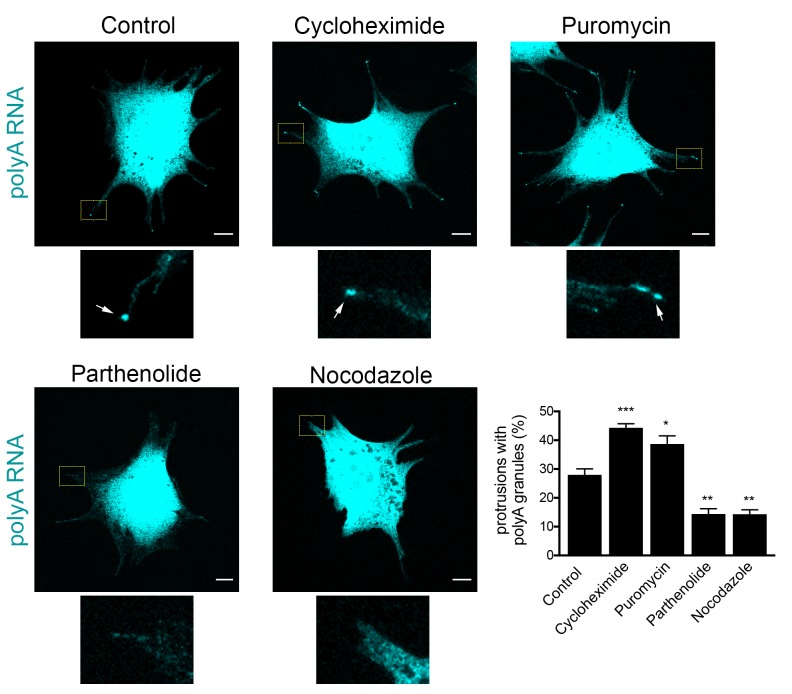
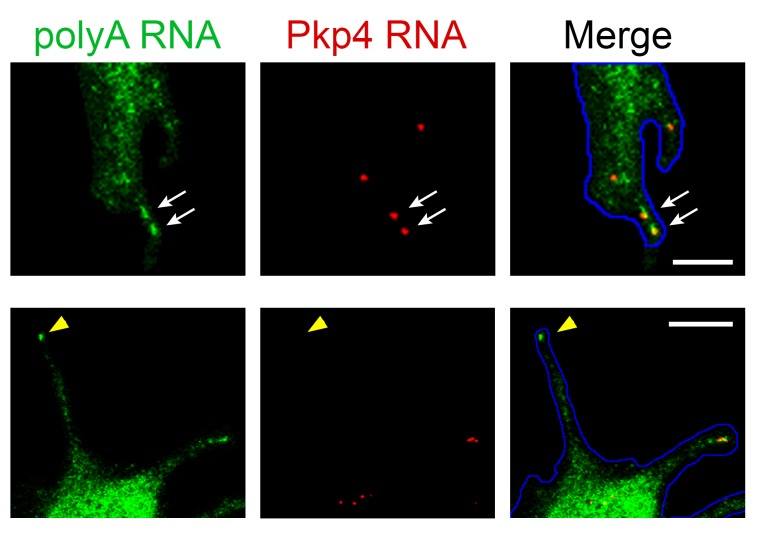
Localization of RNAs to various subcellular destinations is a widely used mechanism that regulates a large proportion of transcripts in polarized cells. In many cases, such localized transcripts mediate spatial control of gene expression by being translationally silent while in transit and locally activated at their destination. Here, we investigate the translation of RNAs localized at dynamic cellular protrusions of human and mouse, migrating, mesenchymal cells. In contrast to the model described above, we find that protrusion-localized RNAs are not locally activated solely at protrusions, but can be translated with similar efficiency in both internal and peripheral locations. Interestingly, protrusion-localized RNAs are translated at extending protrusions, they become translationally silenced in retracting protrusions and this silencing is accompanied by coalescence of single RNAs into larger heterogeneous RNA clusters. This work describes a distinct mode of translational regulation of localized RNAs, which we propose is used to regulate protein activities during dynamic cellular responses.
RNA 在各种亚细胞靶位的定位是一种广泛使用的调控机制,它可以调节大部分极化细胞中的转录本。在许多情况下,这些定位于局部的转录本通过在运输过程中翻译沉默,并在靶位处局部激活,从而介导基因表达的空间调控。在这里,我们研究了定位于人类和小鼠迁移、间充质细胞的动态细胞突起中的 RNA 的翻译。与上述模型相反,我们发现,突起定位的 RNA 不仅在突起处局部激活,而且在内部和外围位置以相似的效率进行翻译。有趣的是,突起定位的 RNA 在延伸的突起中被翻译,它们在回缩的突起中翻译沉默,这种沉默伴随着单个 RNA 聚合成更大的异质 RNA 簇。这项工作描述了一种局部 RNA 翻译调控的独特模式,我们提出这种模式用于在动态细胞反应过程中调节蛋白质活性。