Department of Neurosciences, School of Medicine, Case Western Reserve University, Cleveland, United States.
Elife. 2019 Jul 29;8:e48788. doi: 10.7554/eLife.48788.
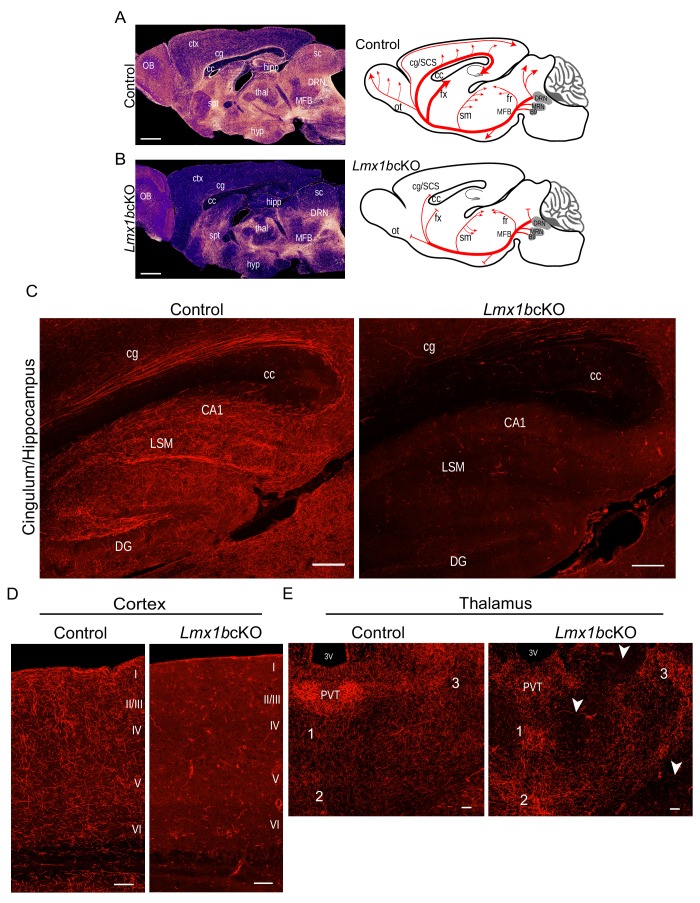
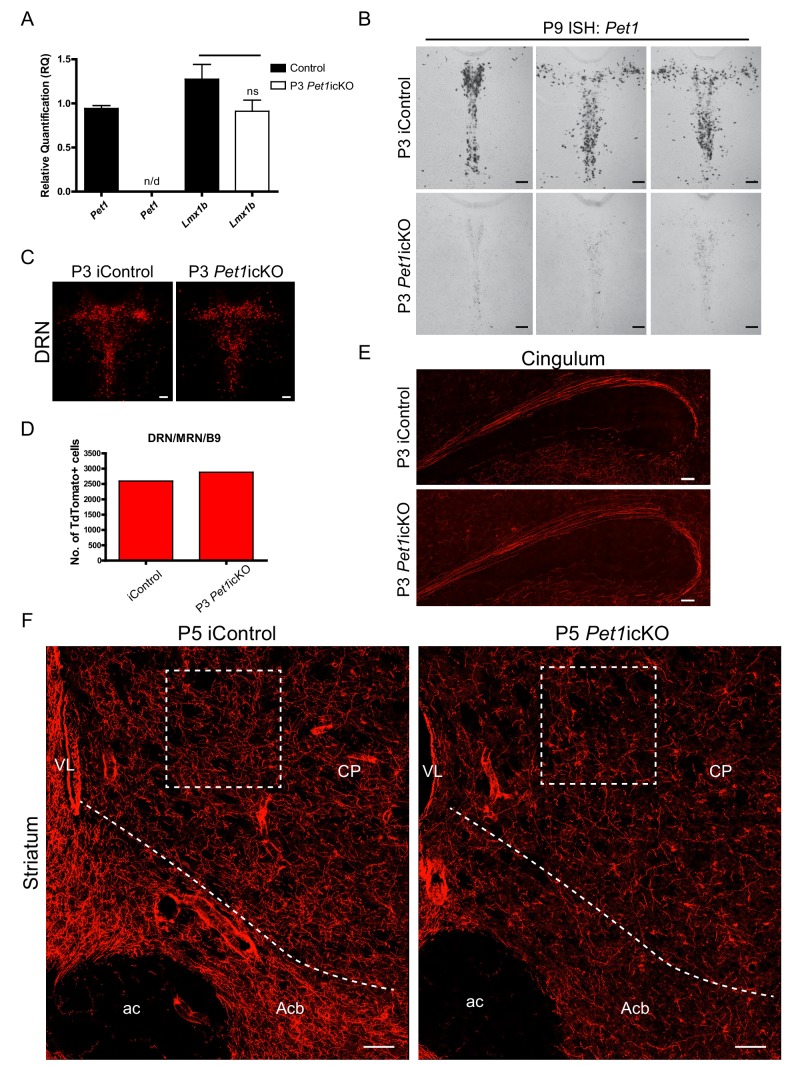
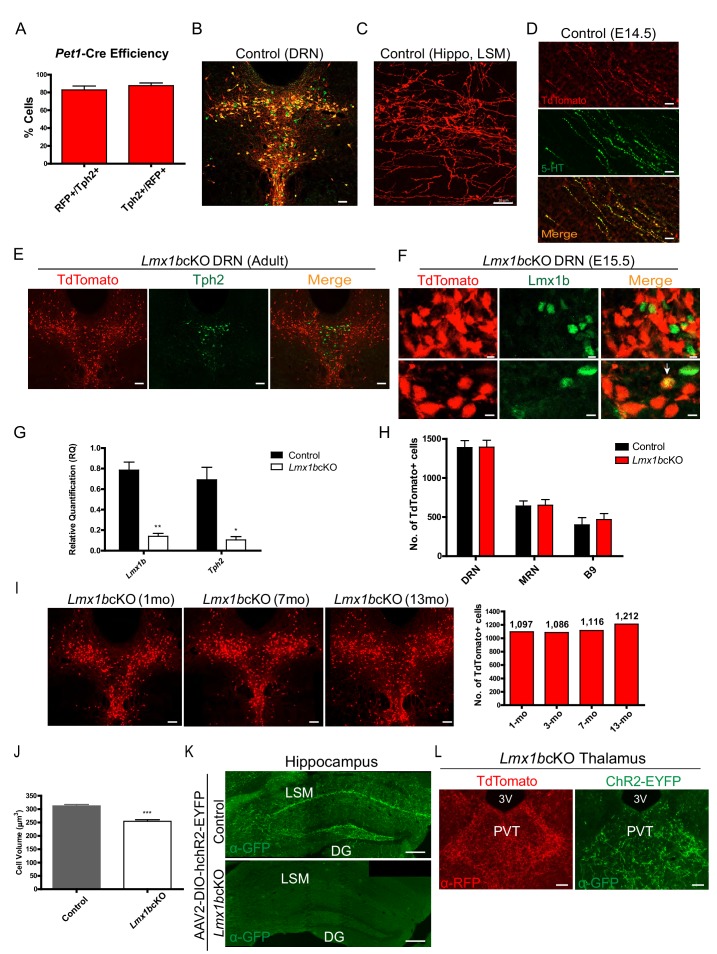
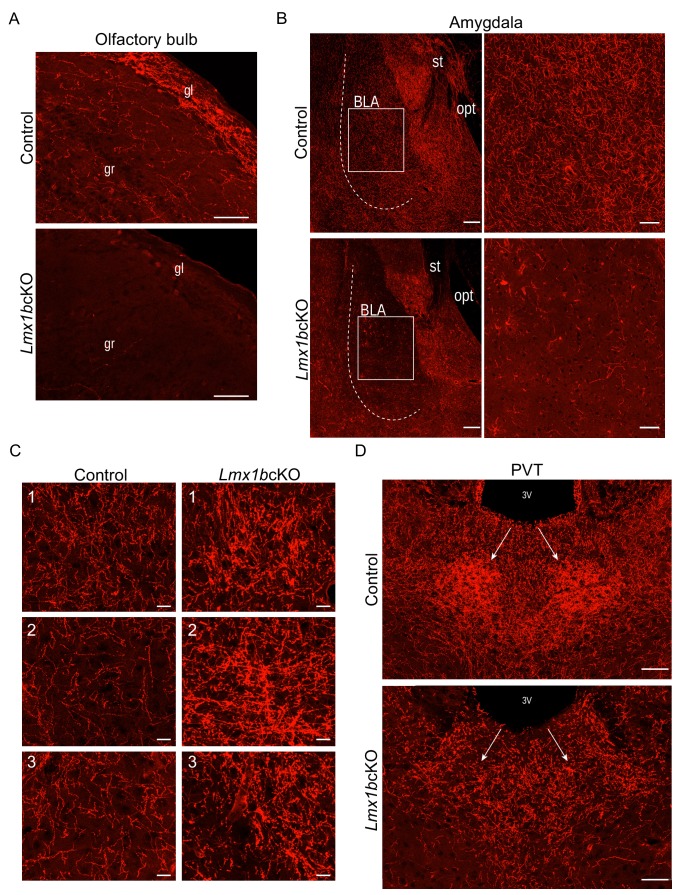
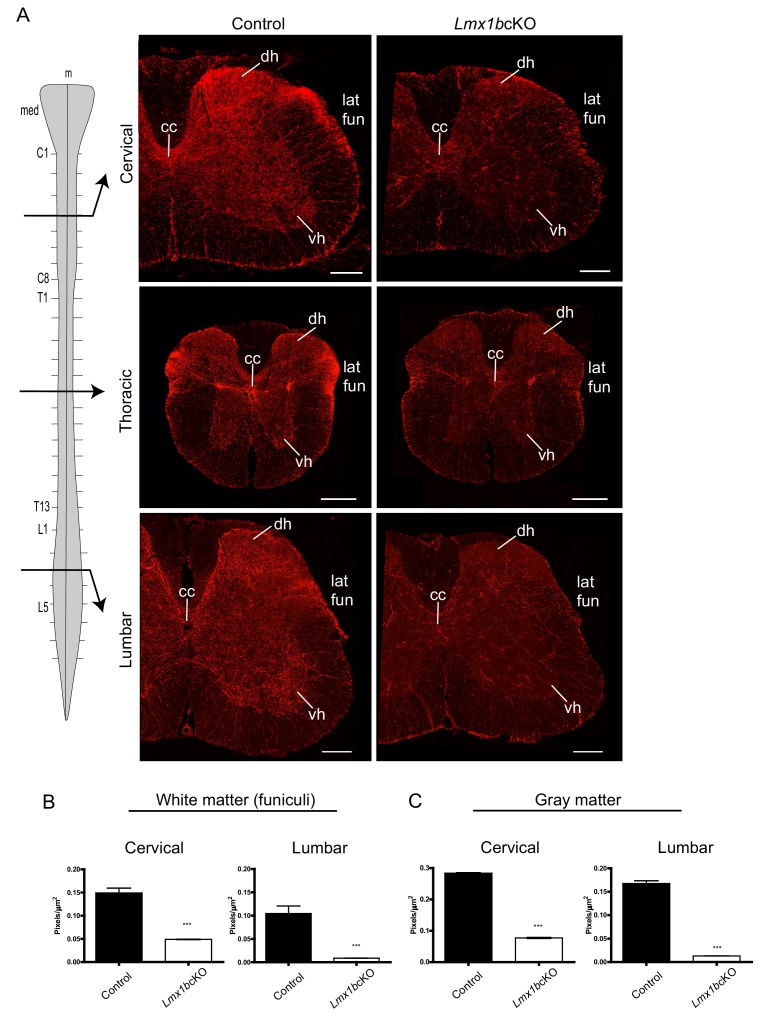
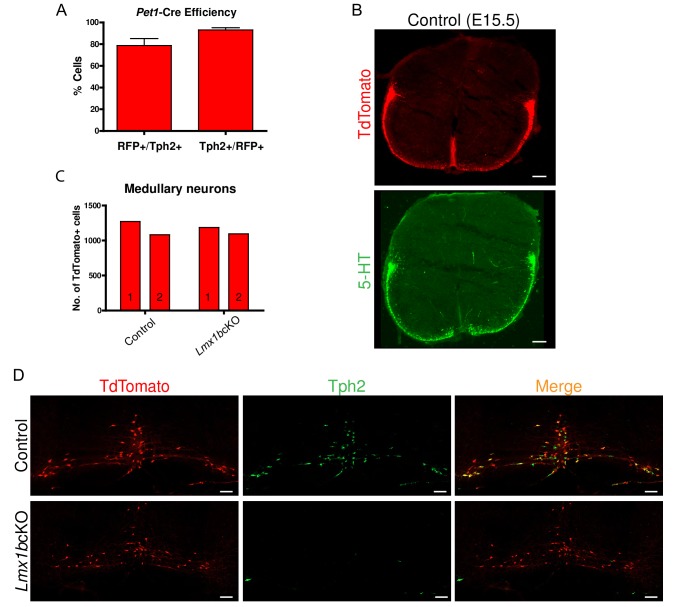
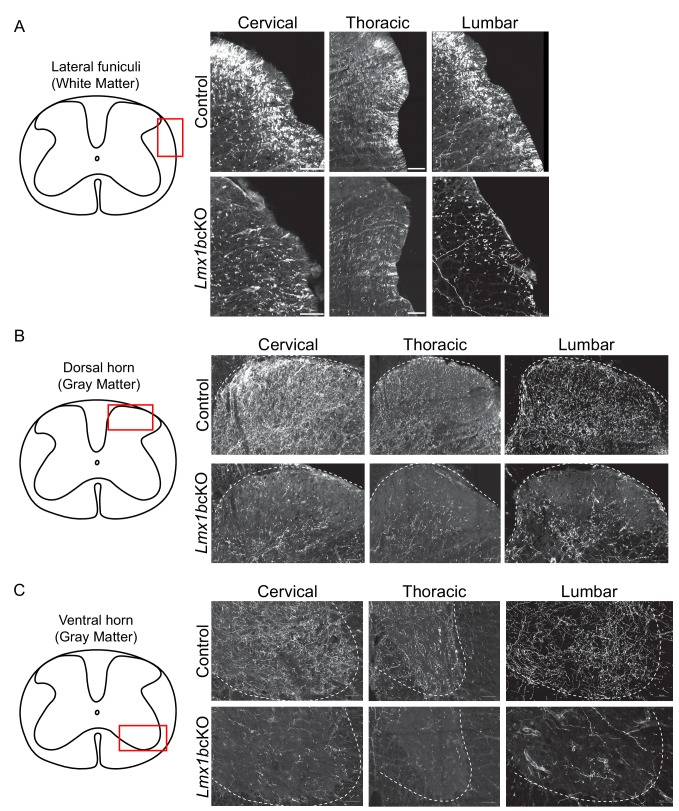
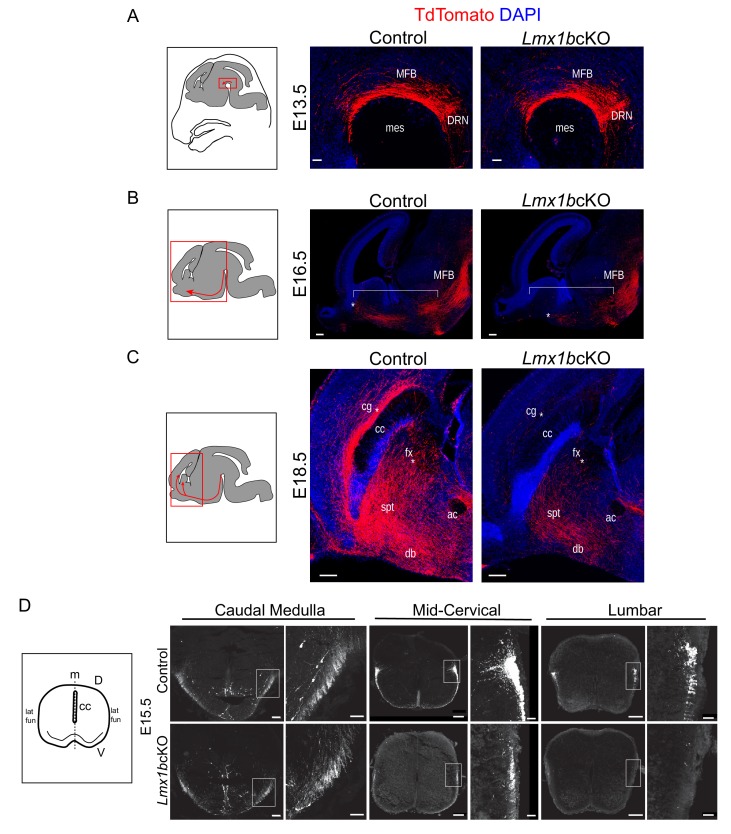
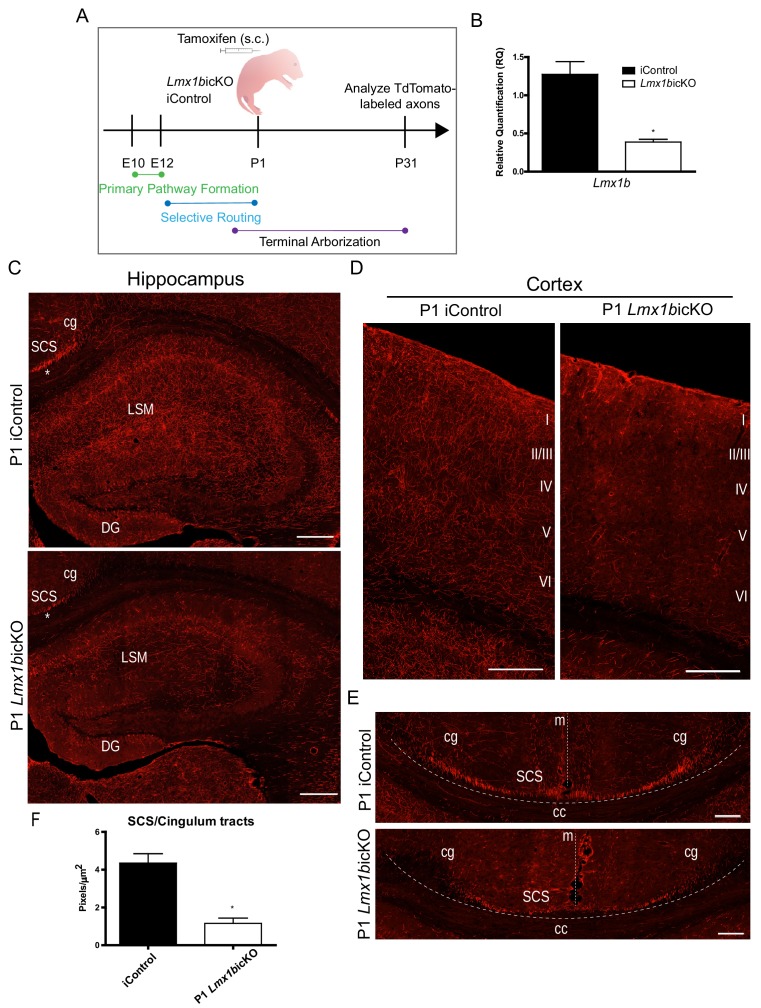
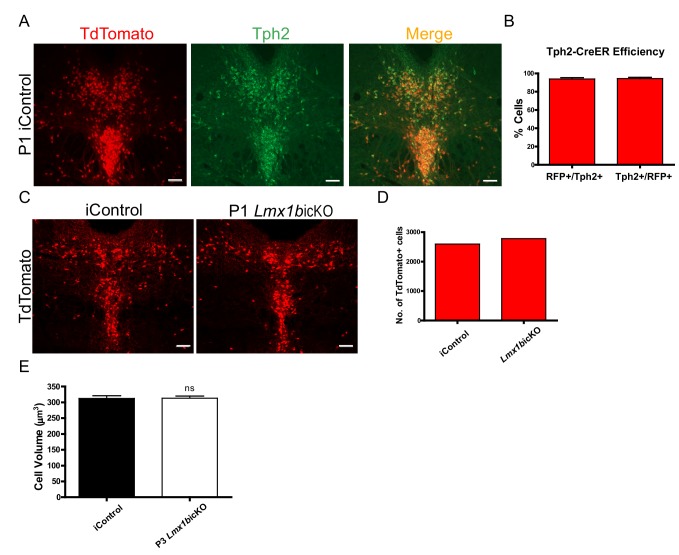
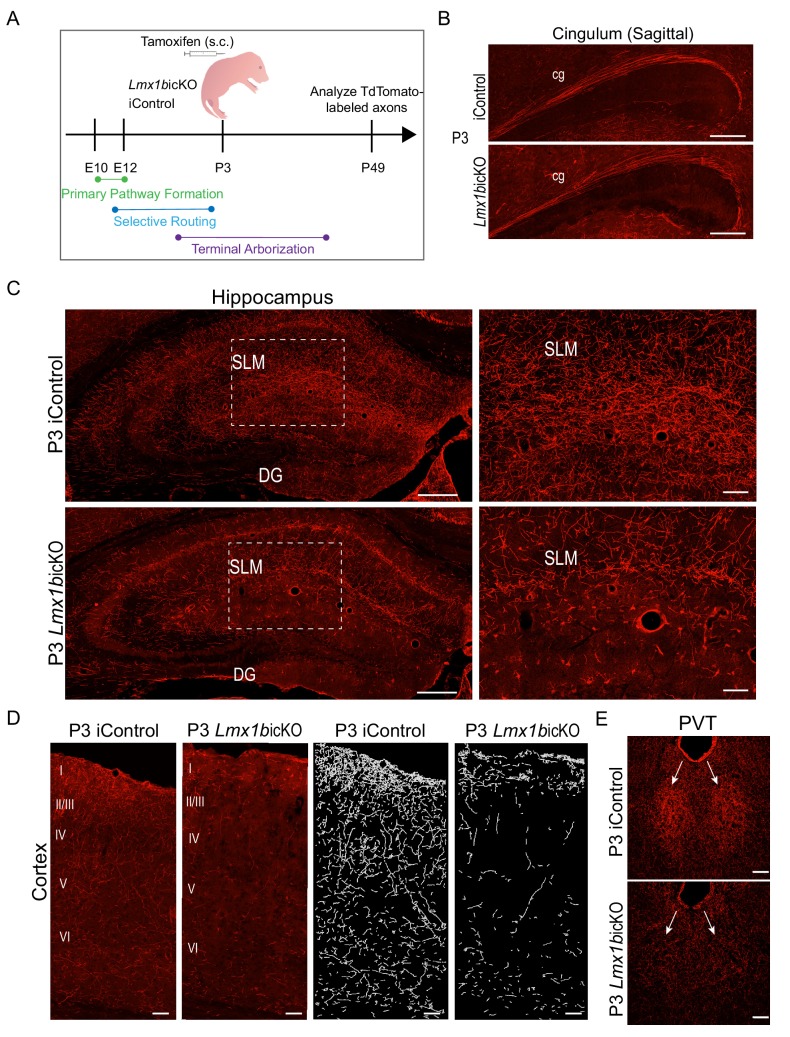
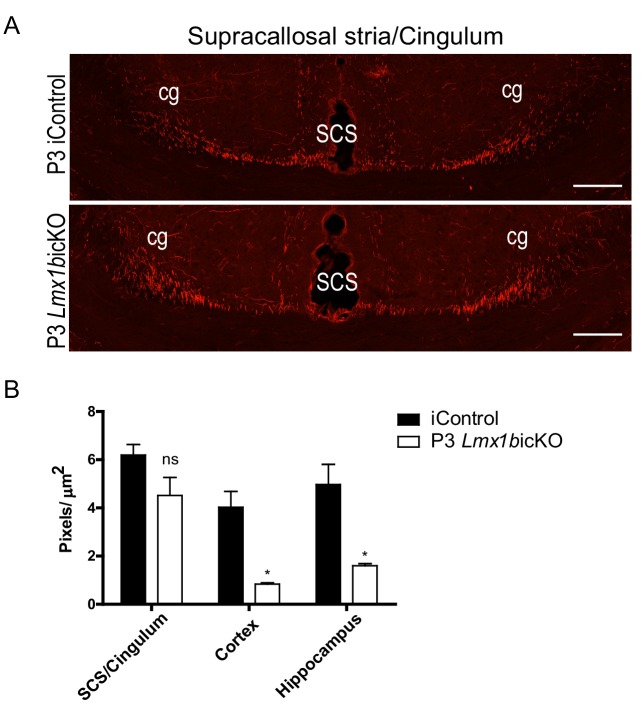
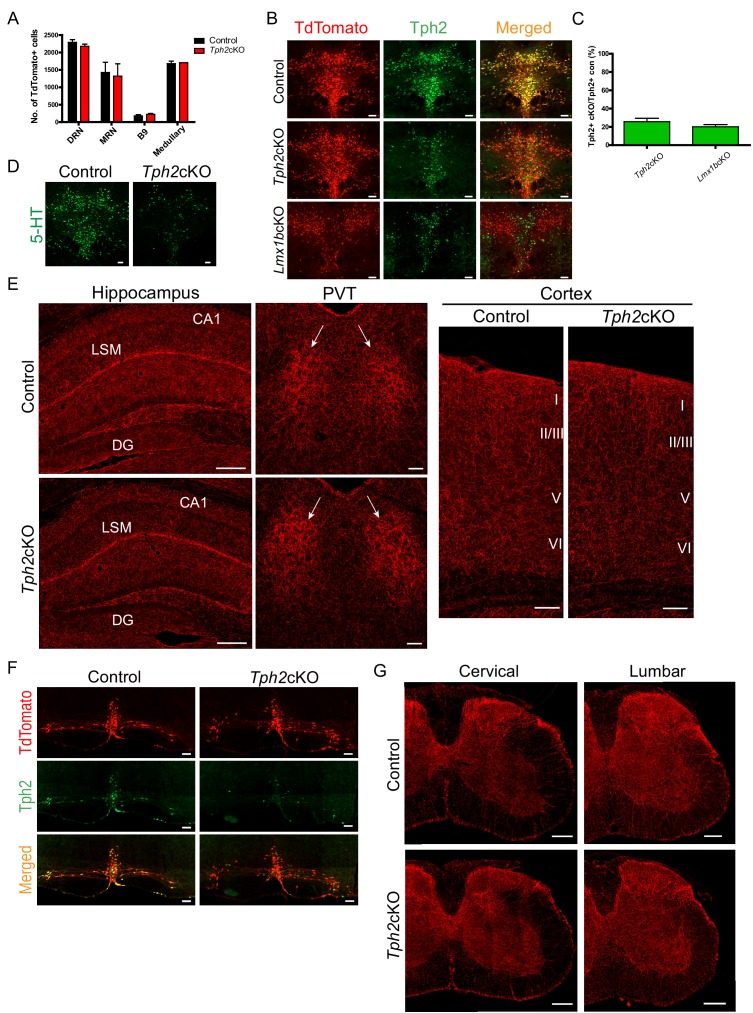
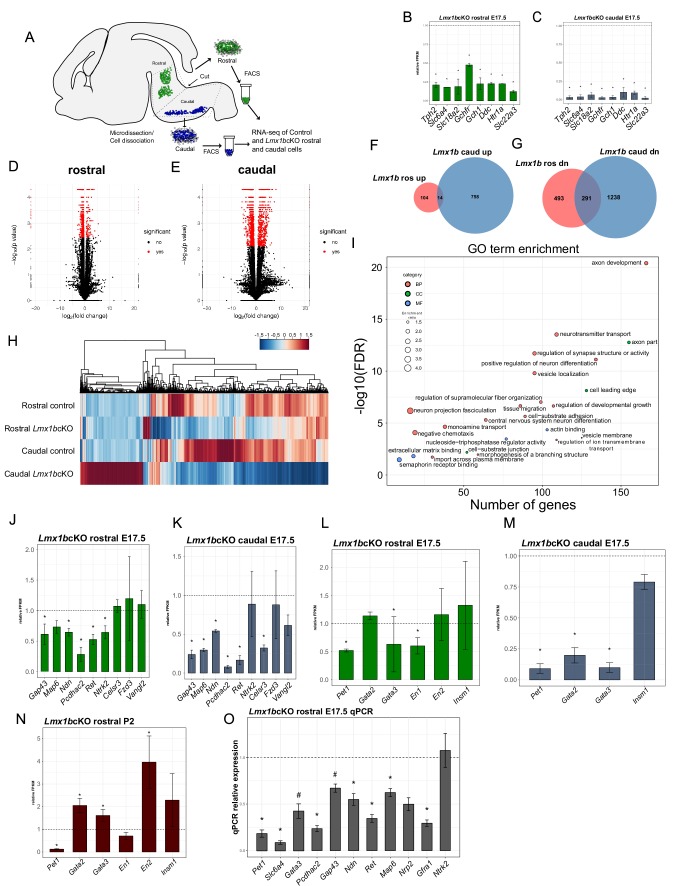
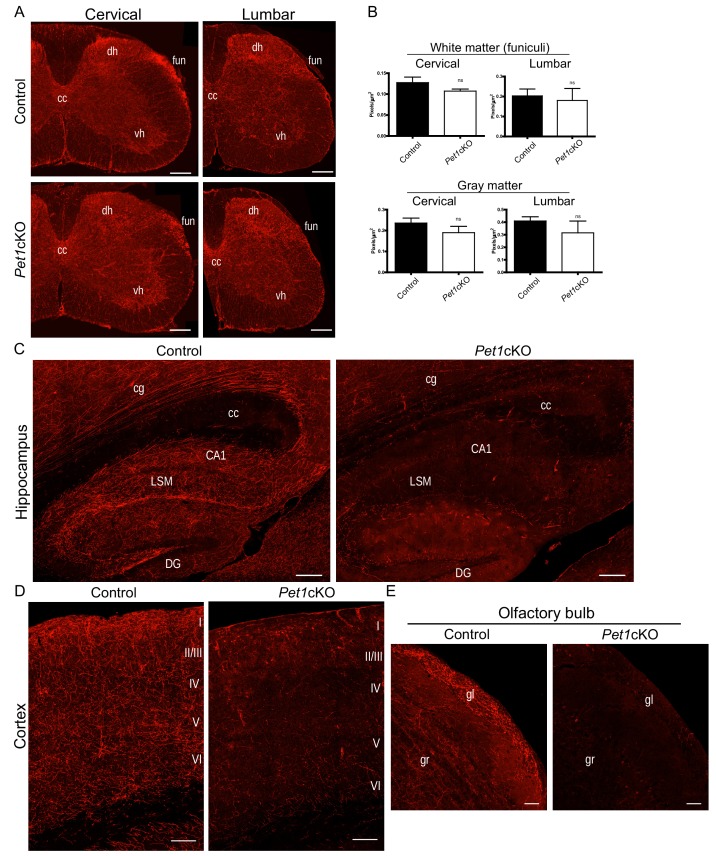
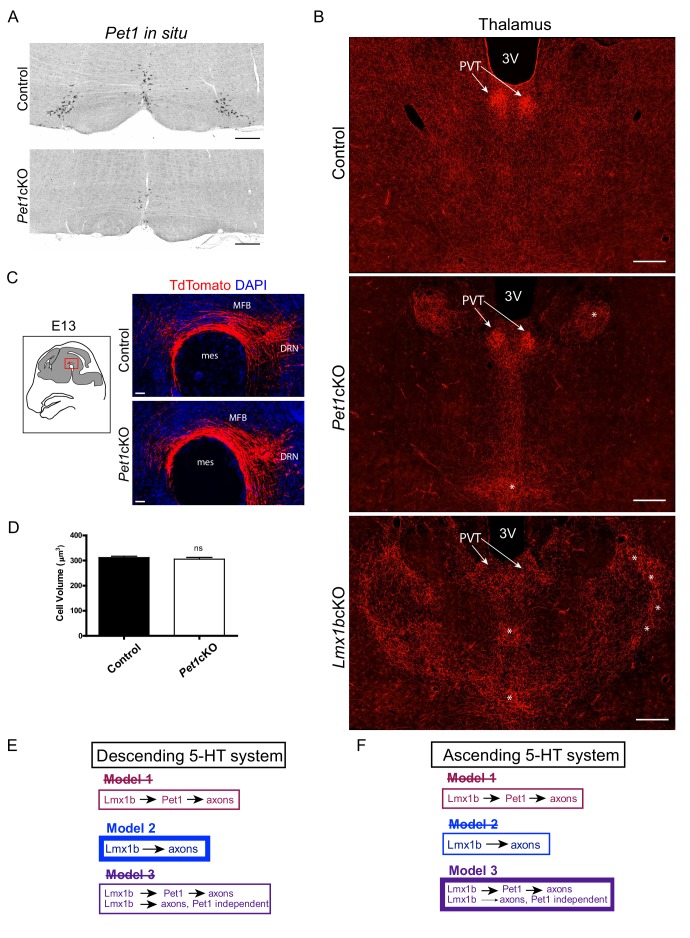
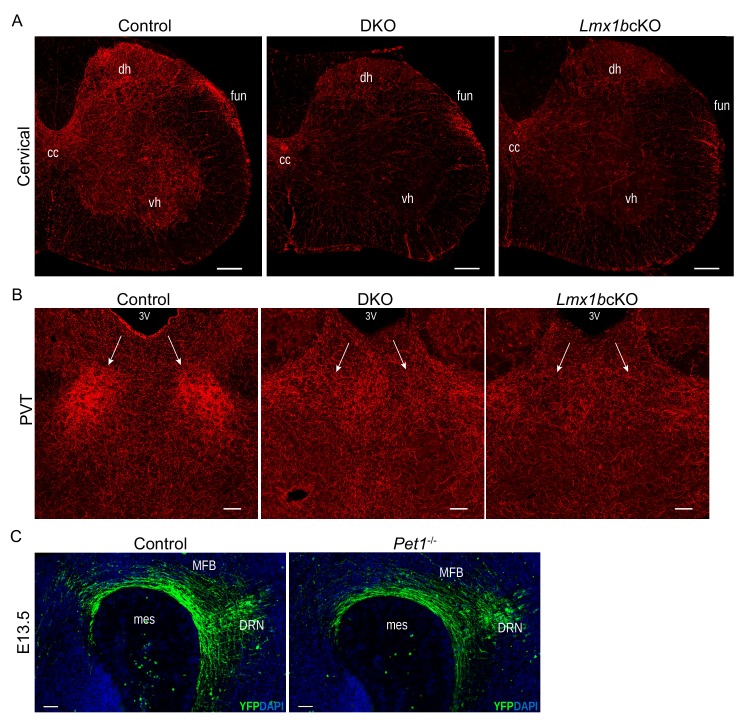
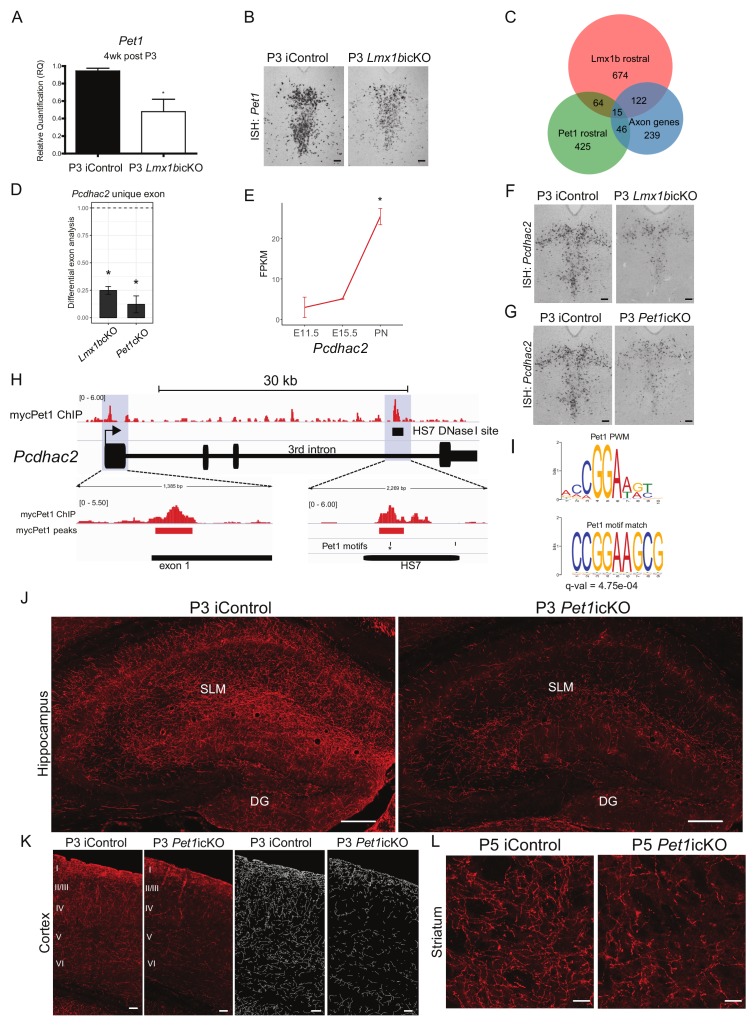
Formation of long-range axons occurs over multiple stages of morphological maturation. However, the intrinsic transcriptional mechanisms that temporally control different stages of axon projection development are unknown. Here, we addressed this question by studying the formation of mouse serotonin (5-HT) axons, the exemplar of long-range profusely arborized axon architectures. We report that LIM homeodomain factor 1b (Lmx1b)-deficient 5-HT neurons fail to generate axonal projections to the forebrain and spinal cord. Stage-specific targeting demonstrates that Lmx1b is required at successive stages to control 5-HT axon primary outgrowth, selective routing, and terminal arborization. We show a Lmx1b→Pet1 regulatory cascade is temporally required for 5-HT arborization and upregulation of the 5-HT axon arborization gene, Protocadherin-alphac2, during postnatal development of forebrain 5-HT axons. Our findings identify a temporal regulatory mechanism in which a single continuously expressed transcription factor functions at successive stages to orchestrate the progressive development of long-range axon architectures enabling expansive neuromodulation.
长程轴突的形成发生在形态成熟的多个阶段。然而,暂时控制轴突投射发育不同阶段的内在转录机制尚不清楚。在这里,我们通过研究小鼠血清素 (5-HT) 轴突的形成来解决这个问题,5-HT 轴突是长程广泛分支轴突结构的范例。我们报告说,LIM 同源框因子 1b (Lmx1b) 缺陷的 5-HT 神经元无法将轴突投射到前脑和脊髓。阶段特异性靶向表明,Lmx1b 在连续阶段需要控制 5-HT 轴突的初级生长、选择性路由和末端分支。我们表明,Lmx1b→Pet1 调节级联在 5-HT 轴突分支和 5-HT 轴突分支基因 Protocadherin-alphac2 的上调的时间上是必需的,这发生在 5-HT 轴突的大脑前脑发育的后期。我们的发现确定了一个时间调节机制,其中单个连续表达的转录因子在连续阶段发挥作用,以协调长程轴突结构的渐进发展,从而实现广泛的神经调制。