Institute of Biochemistry, ETH Zürich, Zurich, Switzerland.
Department of Biosystems Science and Engineering, ETH Zürich, SIB Swiss Institute of Bioinformatics, Basel, Switzerland.
Elife. 2019 Sep 6;8:e48627. doi: 10.7554/eLife.48627.
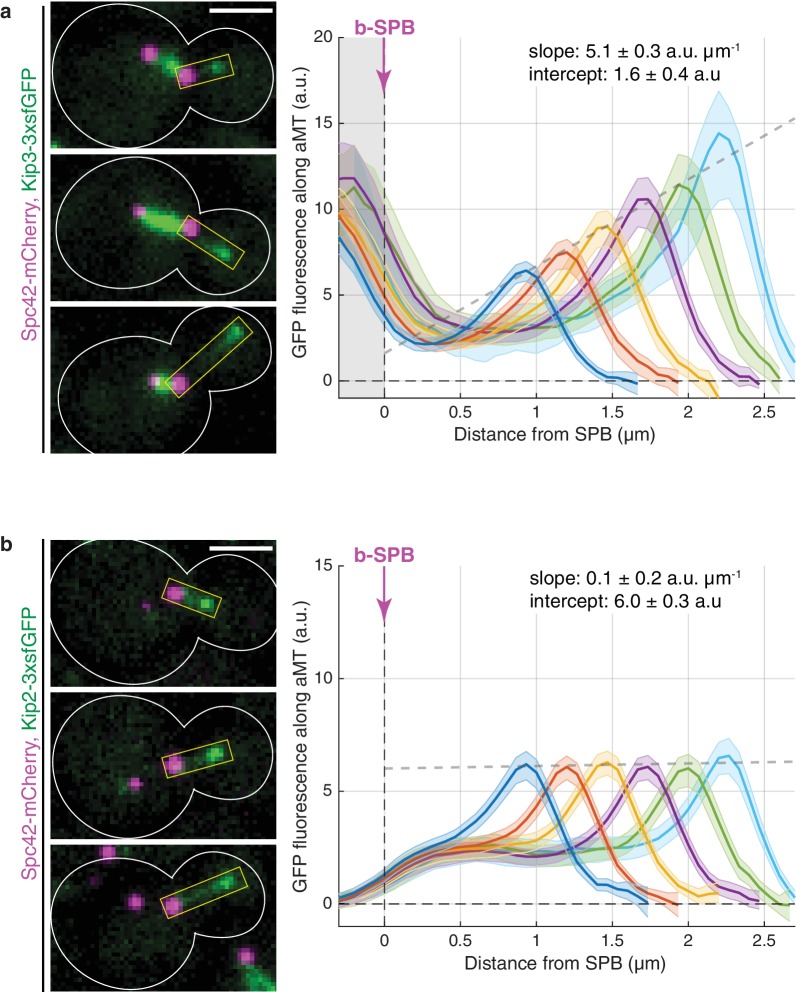
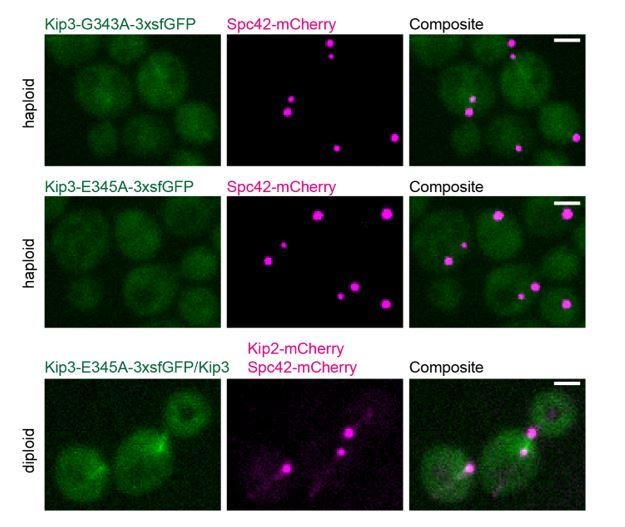
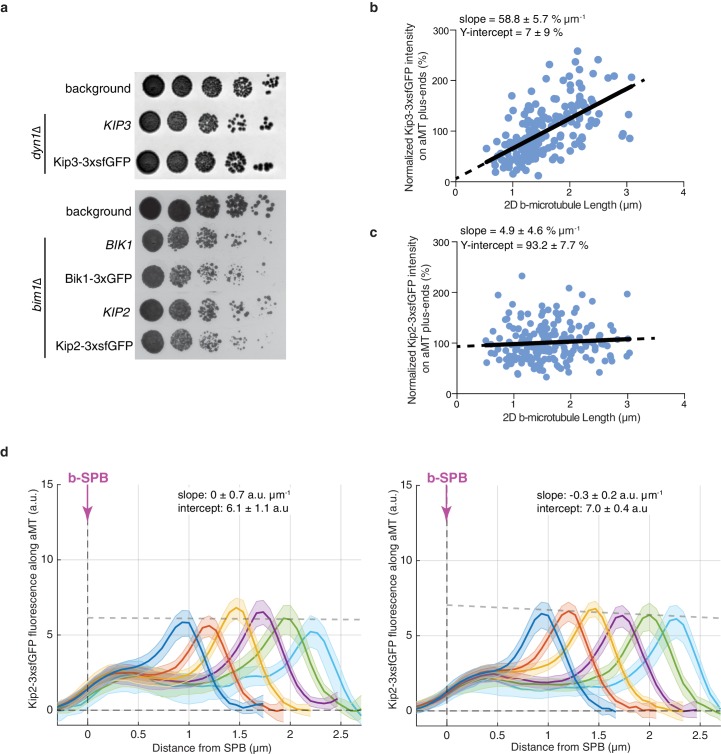
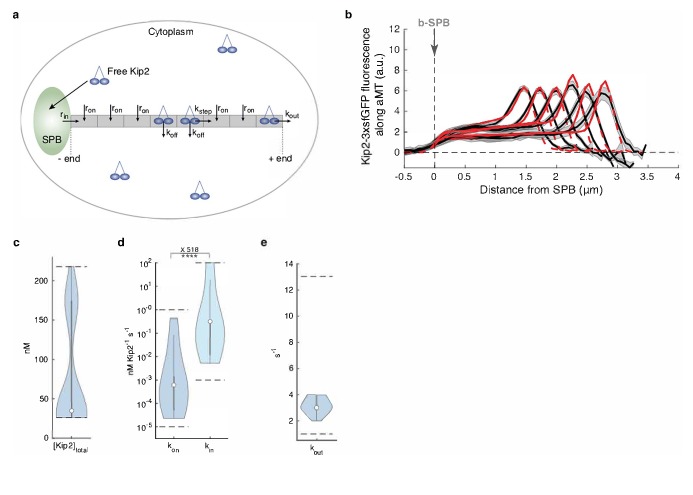
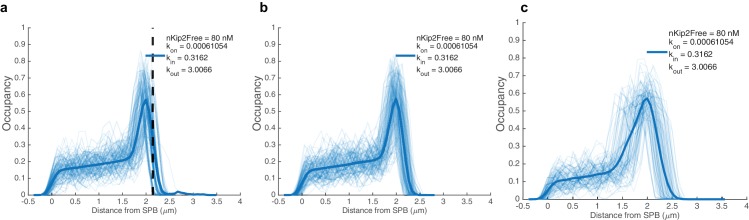
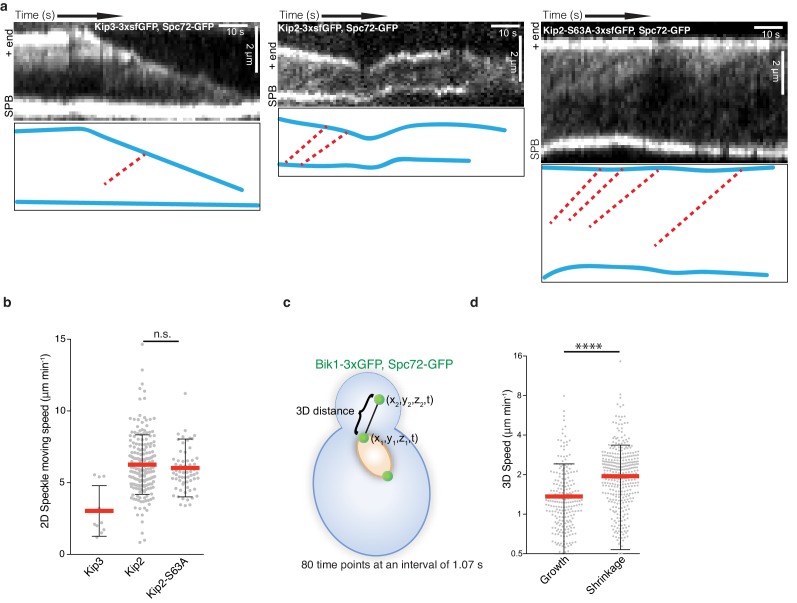
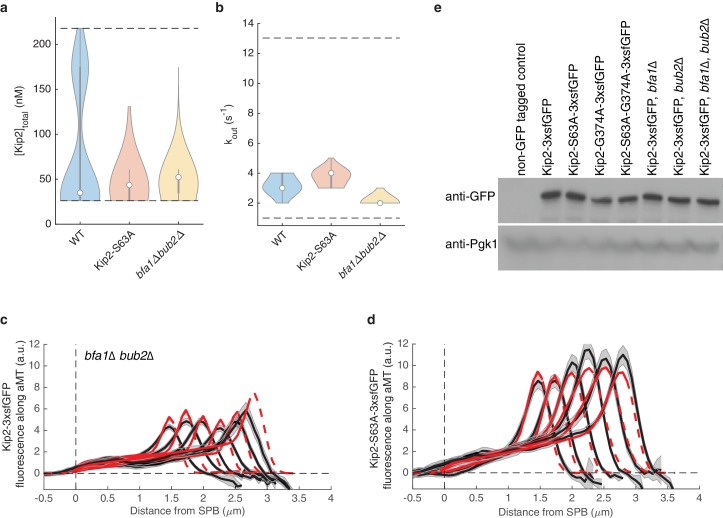
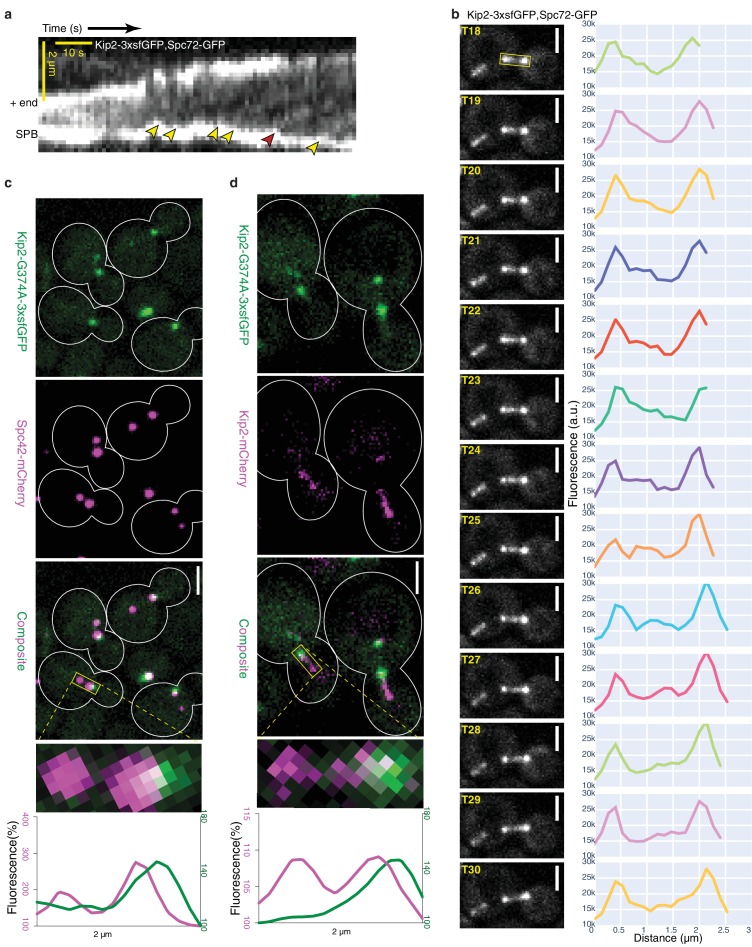
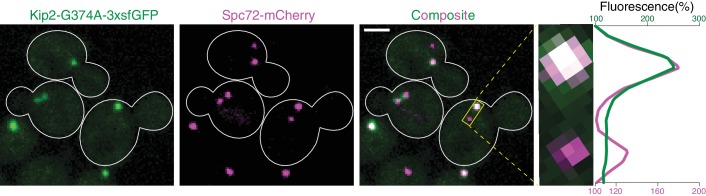
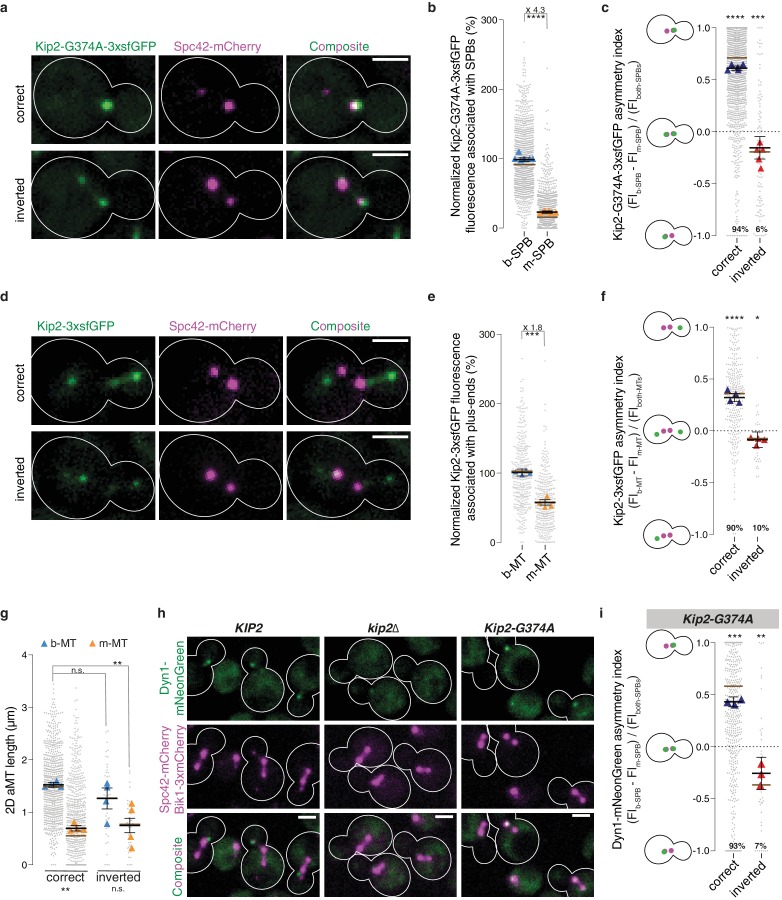
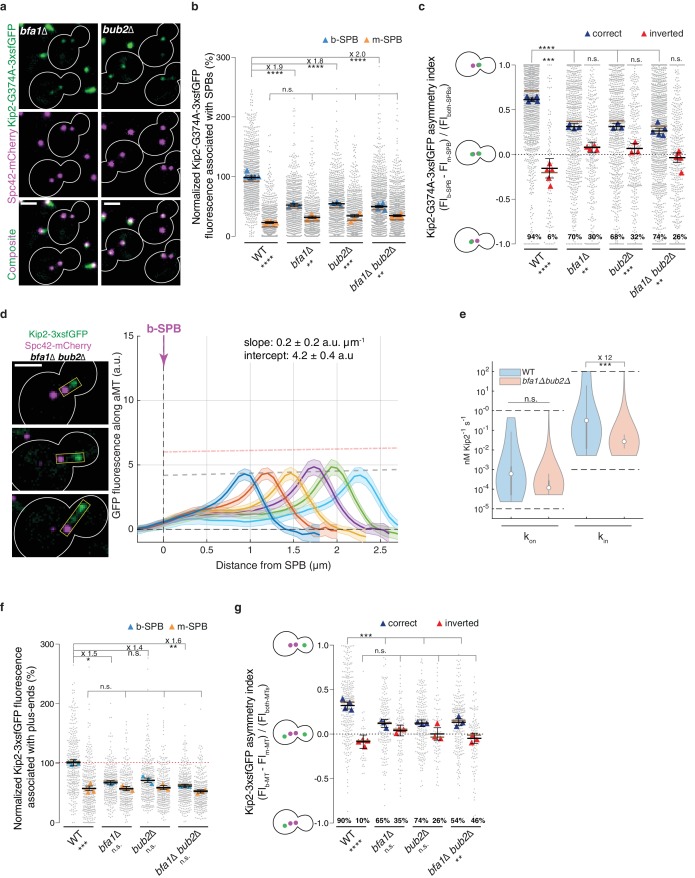
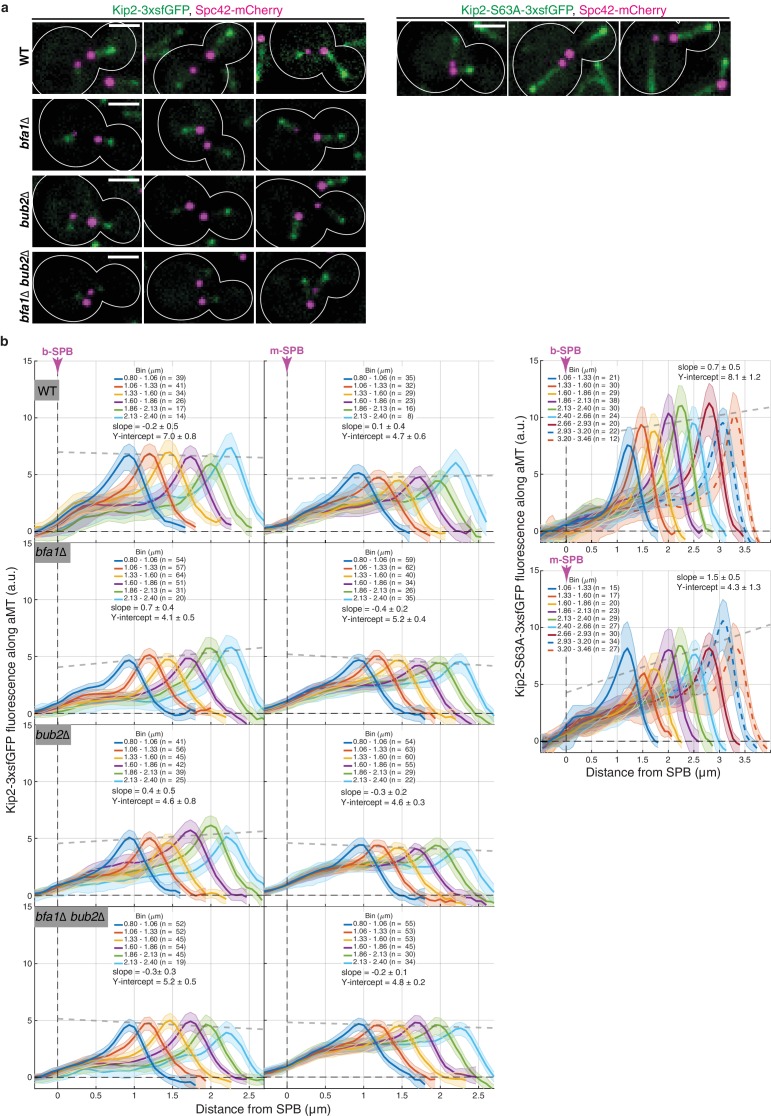
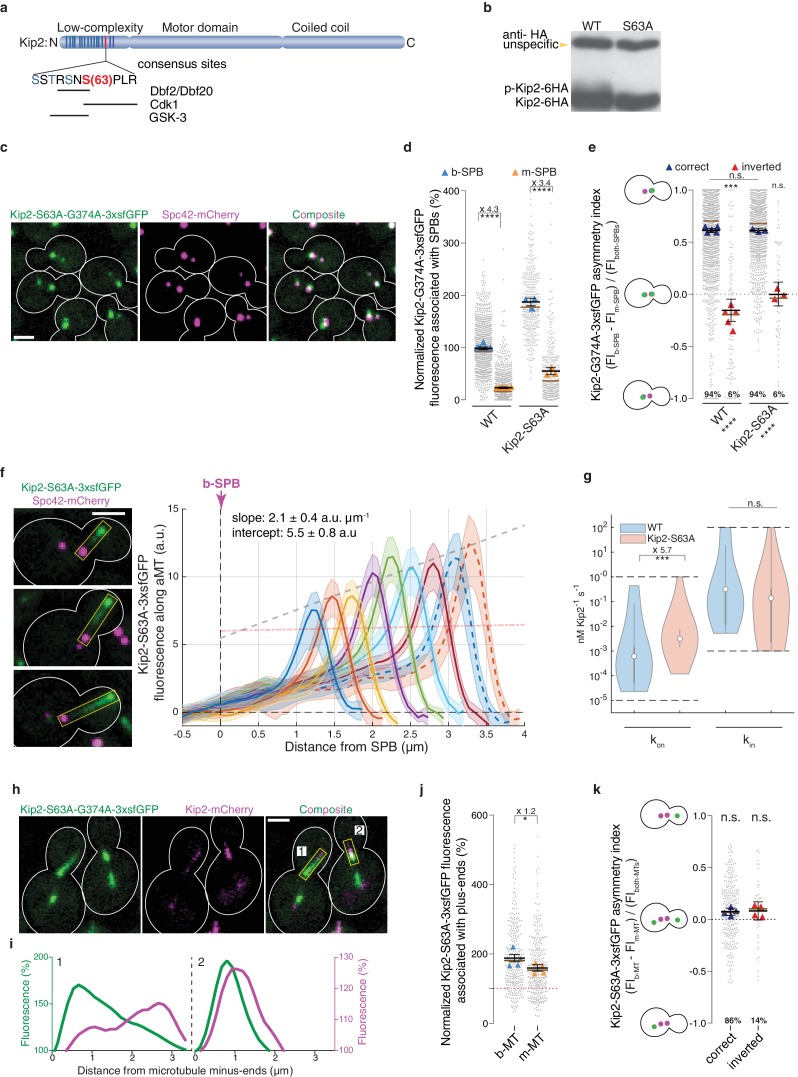
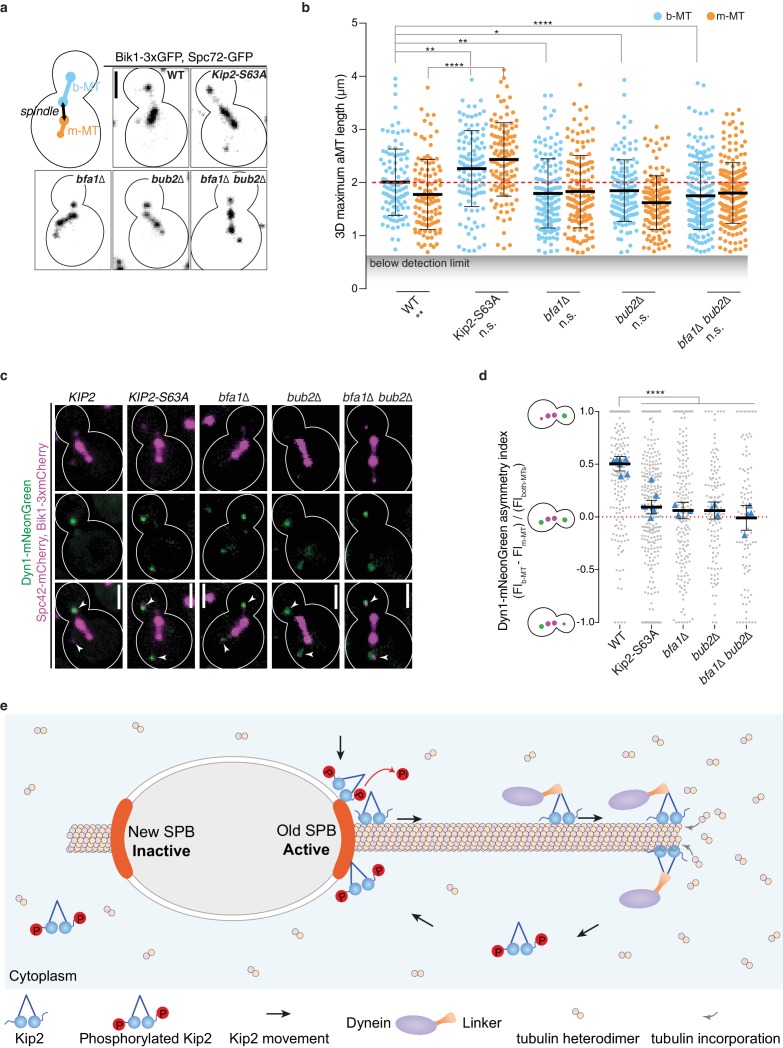
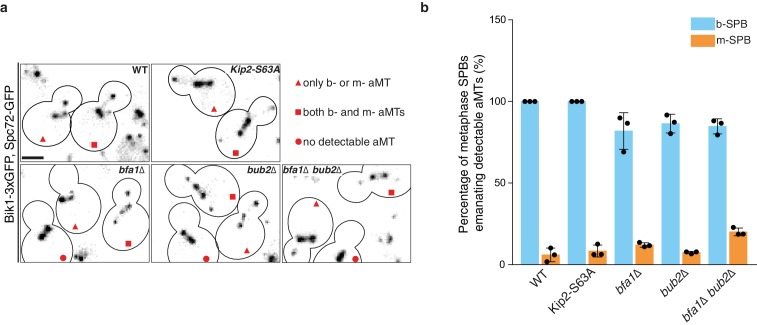
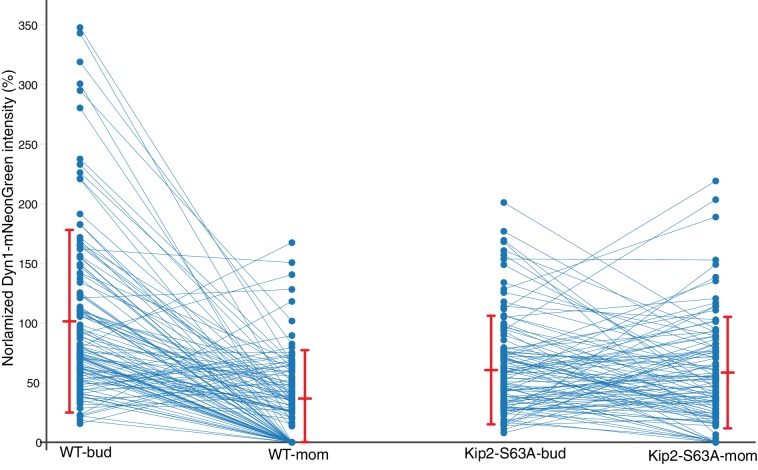
In eukaryotes, the organization and function of the microtubule cytoskeleton depend on the allocation of different roles to individual microtubules. For example, many asymmetrically dividing cells differentially specify microtubule behavior at old and new centrosomes. Here we show that yeast spindle pole bodies (SPBs, yeast centrosomes) differentially control the plus-end dynamics and cargoes of their astral microtubules, remotely from the minus-end. The old SPB recruits the kinesin motor protein Kip2, which then translocates to the plus-end of the emanating microtubules, promotes their extension and delivers dynein into the bud. Kip2 recruitment at the SPB depends on Bub2 and Bfa1, and phosphorylation of cytoplasmic Kip2 prevents random lattice binding. Releasing Kip2 of its control by SPBs equalizes its distribution, the length of microtubules and dynein distribution between the mother cell and its bud. These observations reveal that microtubule organizing centers use minus to plus-end directed remote control to individualize microtubule function.
在真核生物中,微管细胞骨架的组织和功能依赖于将不同的角色分配给各个微管。例如,许多不对称分裂的细胞在旧的和新的中心体处特异性地指定微管行为。在这里,我们表明酵母纺锤体极体(SPB,酵母中心体)从负端远程控制其星体微管的正端动力学和货物。旧的 SPB 招募了驱动蛋白 Kip2 运动蛋白,然后将其转移到发出的微管的正端,促进其延伸并将动力蛋白输送到芽中。SPB 处的 Kip2 募集依赖于 Bub2 和 Bfa1,细胞质 Kip2 的磷酸化防止随机格子结合。通过 SPB 释放 Kip2 的控制,使其在母细胞与其芽之间的微管长度和动力蛋白分布均匀化。这些观察结果表明,微管组织中心使用从负端到正端的远程控制来使微管功能个性化。