School of Biological Sciences, Georgia Institute of Technology, Atlanta, United States.
Interdisciplinary Graduate Program in Quantitative Biosciences, Georgia Institute of Technology, Atlanta, United States.
Elife. 2019 Sep 9;8:e48119. doi: 10.7554/eLife.48119.
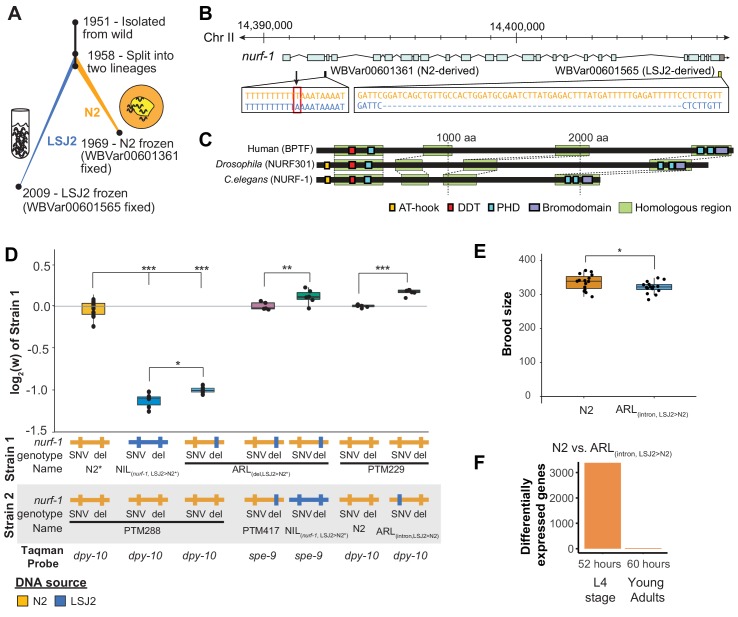
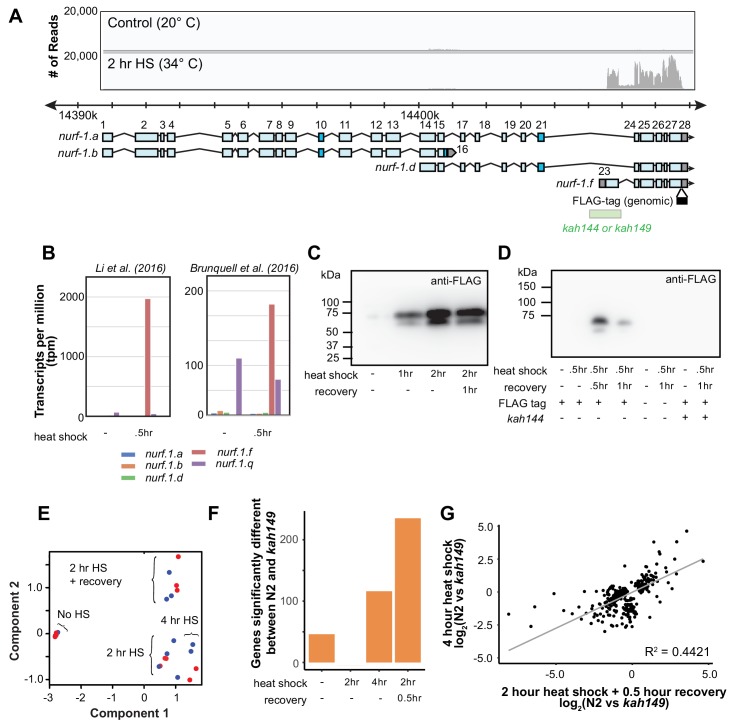
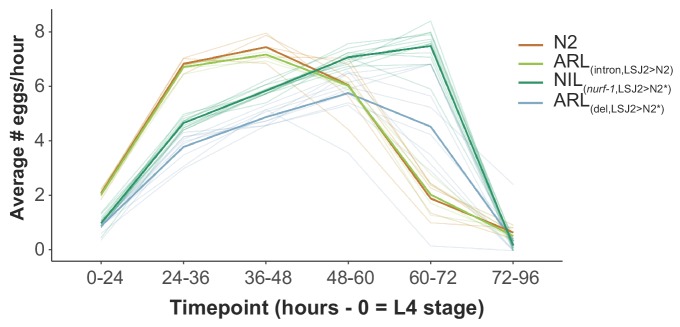
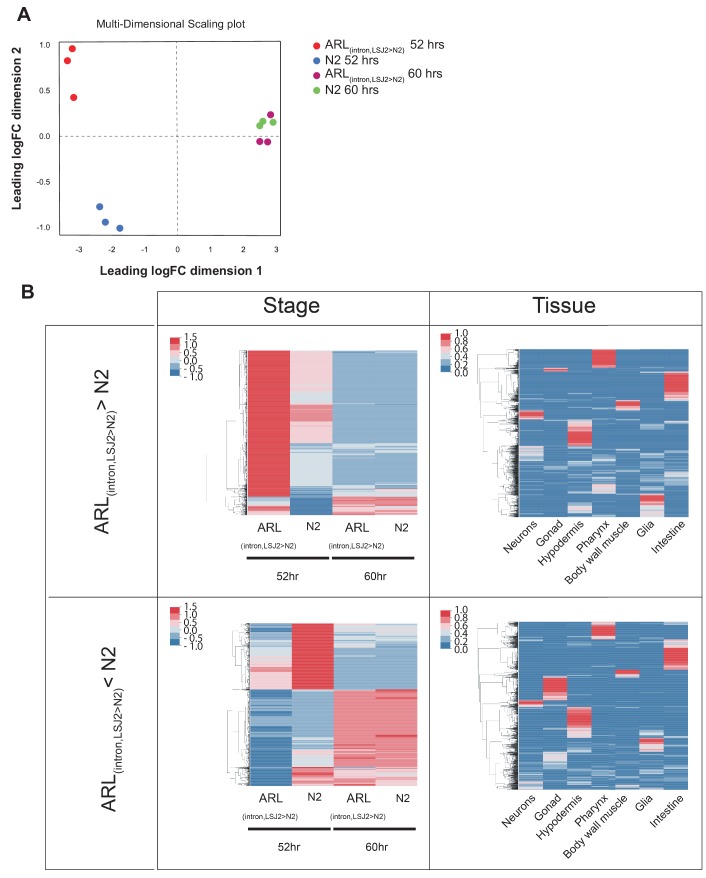
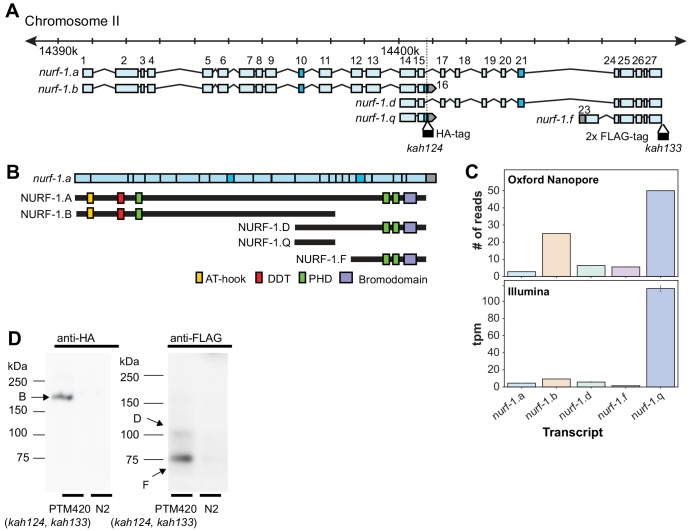
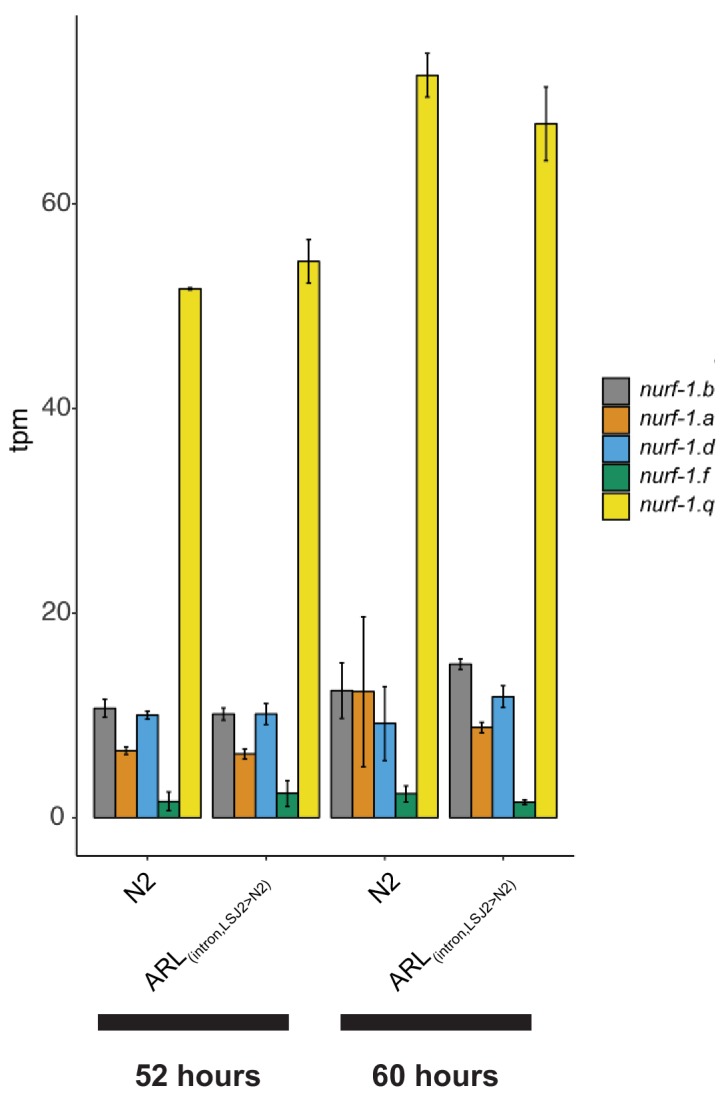
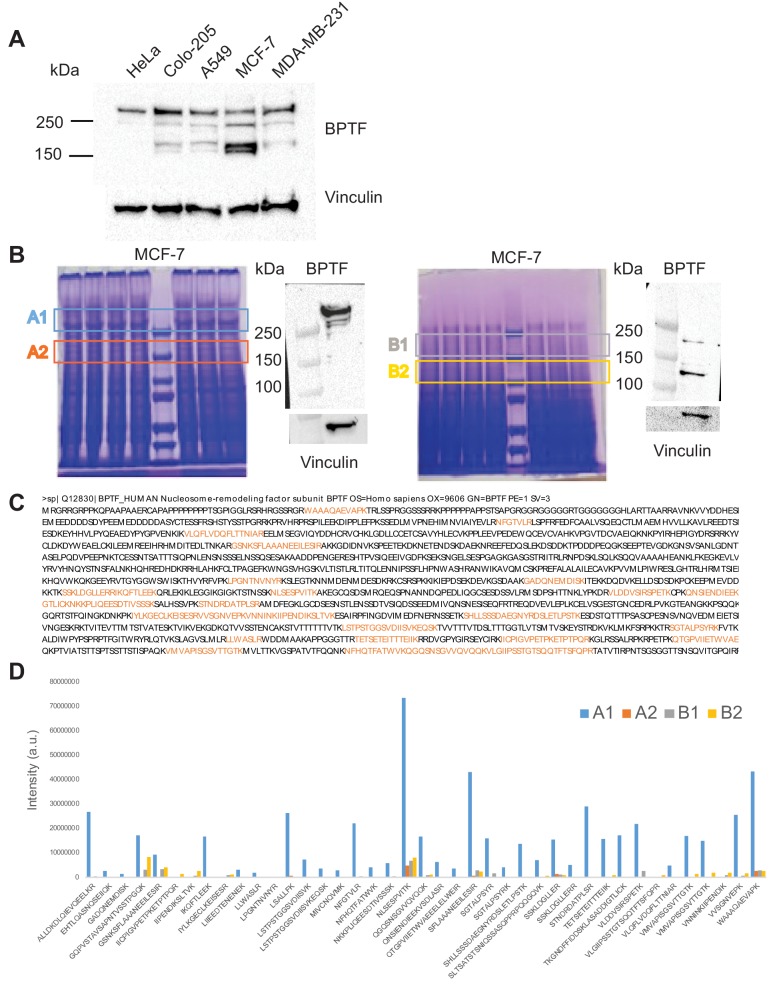
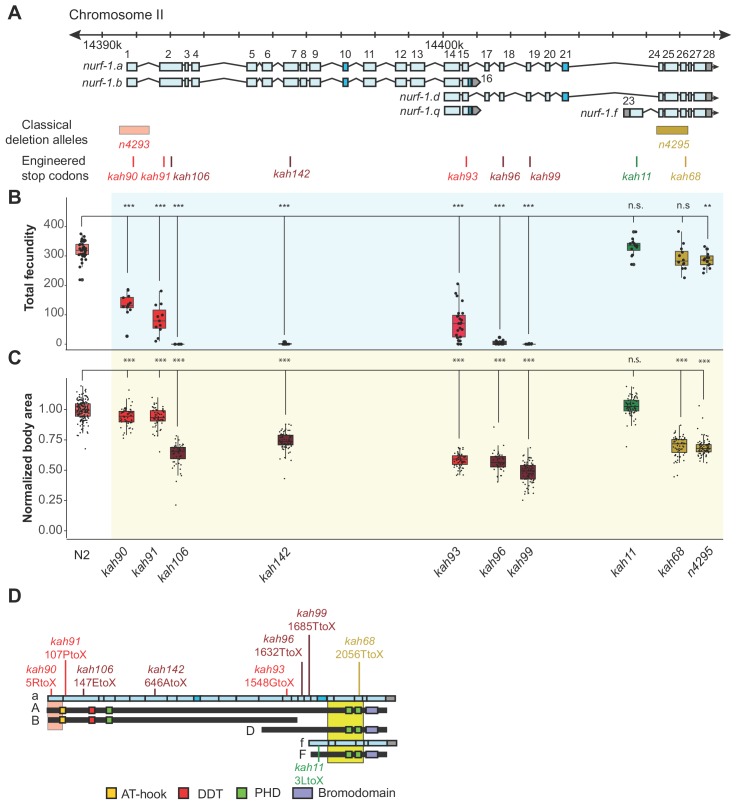
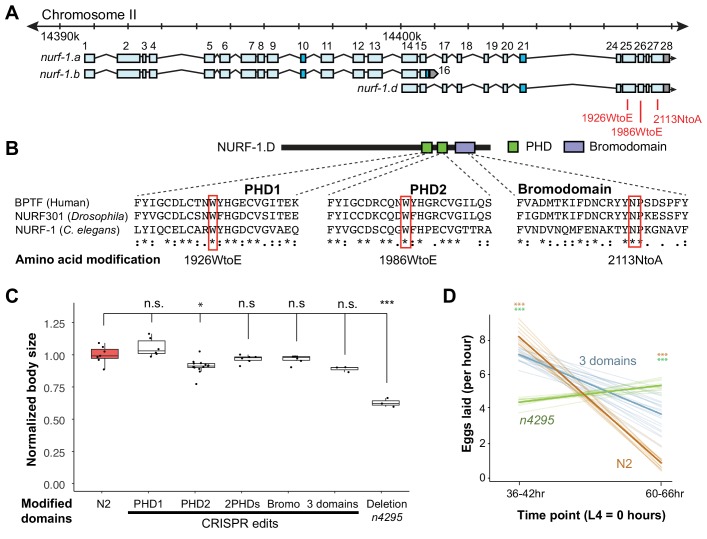
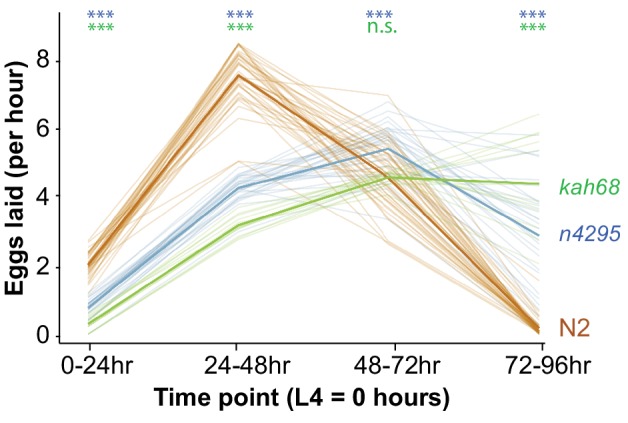
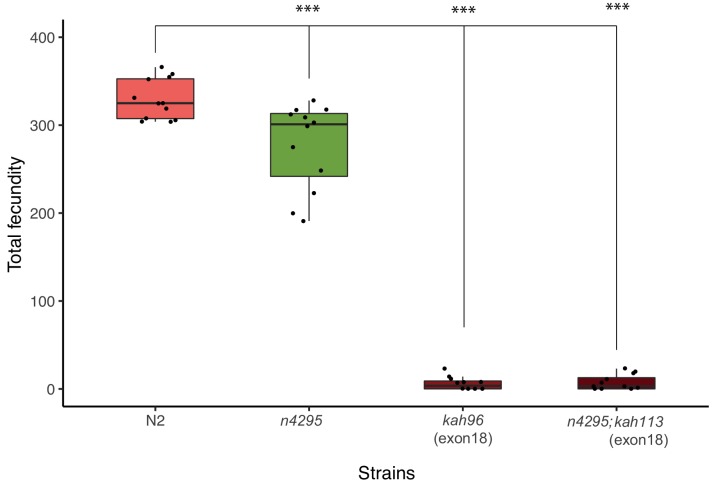
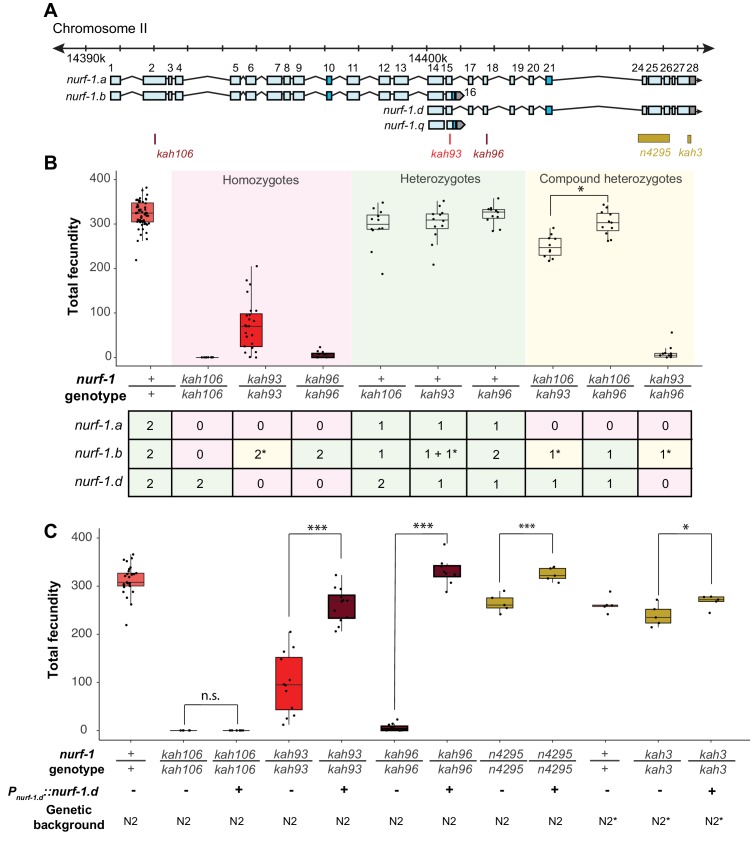
Genes can encode multiple isoforms, broadening their functions and providing a molecular substrate to evolve phenotypic diversity. Evolution of isoform function is a potential route to adapt to new environments. Here we show that de novo, beneficial alleles in the gene became fixed in two laboratory lineages of after isolation from the wild in 1951, before methods of cryopreservation were developed. encodes an ortholog of BPTF, a large (>300 kD) multidomain subunit of the NURF chromatin remodeling complex. Using CRISPR-Cas9 genome editing and transgenic rescue, we demonstrate that in , has split into two, largely non-overlapping isoforms (NURF-1.D and NURF-1.B, which we call Yin and Yang, respectively) that share only two of 26 exons. Both isoforms are essential for normal gametogenesis but have opposite effects on male/female gamete differentiation. Reproduction in hermaphrodites, which involves production of both sperm and oocytes, requires a balance of these opposing Yin and Yang isoforms. Transgenic rescue and genetic position of the fixed mutations suggest that different isoforms are modified in each laboratory strain. In a related clade of nematodes, the shared exons have duplicated, resulting in the split of the Yin and Yang isoforms into separate genes, each containing approximately 200 amino acids of duplicated sequence that has undergone accelerated protein evolution following the duplication. Associated with this duplication event is the loss of two additional transcripts, including the long-form transcript and a newly identified, highly expressed transcript encoded by the duplicated exons. We propose these lost transcripts are non-functional side products necessary to transcribe the Yin and Yang transcripts in the same cells. Our work demonstrates how gene sharing, through the production of multiple isoforms, can precede the creation of new, independent genes.
基因可以编码多个异构体,从而拓宽其功能,并为进化表型多样性提供分子基础。异构体功能的进化是适应新环境的潜在途径。在这里,我们表明,在 1951 年从野外分离后,在开发冷冻保存方法之前,基因中的新有益等位基因在两个实验室谱系中固定下来。 编码 BPTF 的同源物,NURF 染色质重塑复合物的一个大(>300kD)多结构域亚基。使用 CRISPR-Cas9 基因组编辑和转基因拯救,我们证明,在 中, 已分裂为两个,大部分不重叠的异构体(NURF-1.D 和 NURF-1.B,我们分别称为 Yin 和 Yang),它们仅共享 26 个外显子中的两个。这两个异构体对于正常配子发生都是必需的,但对雄性/雌性配子分化有相反的影响。涉及精子和卵子产生的雌雄同体的繁殖需要这些相反的 Yin 和 Yang 异构体之间的平衡。转基因拯救和固定突变的遗传位置表明,不同的异构体在每个实验室菌株中都发生了修饰。在相关的线虫分支中,共享的外显子已发生重复,导致 Yin 和 Yang 异构体分裂成单独的基因,每个基因都包含大约 200 个氨基酸的重复序列,该序列在重复后经历了加速的蛋白质进化。与这个重复事件相关的是另外两个 转录物的丢失,包括长形式转录物和新鉴定的、高度表达的转录物,由重复的外显子编码。我们提出这些丢失的转录物是不必要的非功能副产物,需要在同一细胞中转录 Yin 和 Yang 转录物。我们的工作表明,通过产生多个异构体来共享基因如何先于新的、独立基因的产生。