The Sir William Dunn School of Pathology, University of Oxford, Oxford, United Kingdom.
Elife. 2019 Sep 9;8:e50130. doi: 10.7554/eLife.50130.
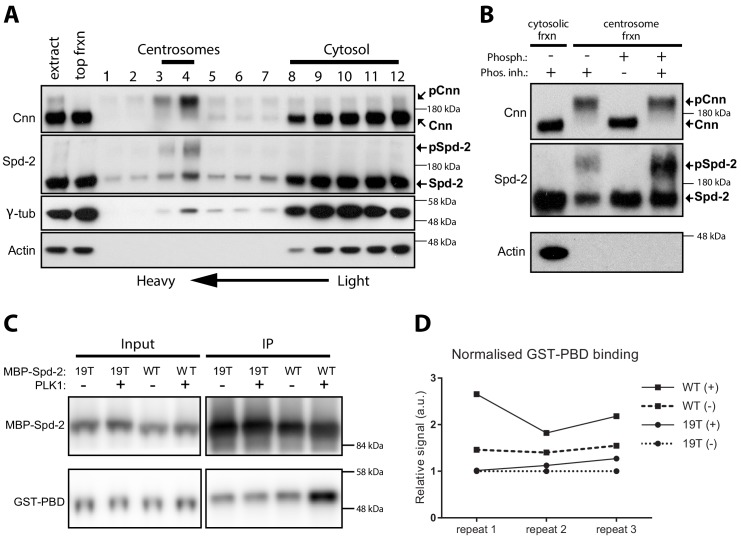
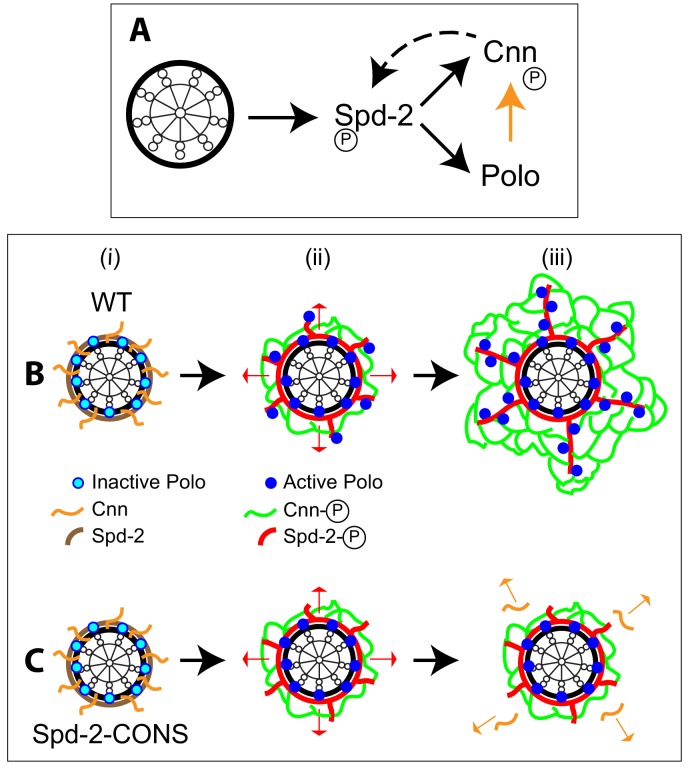
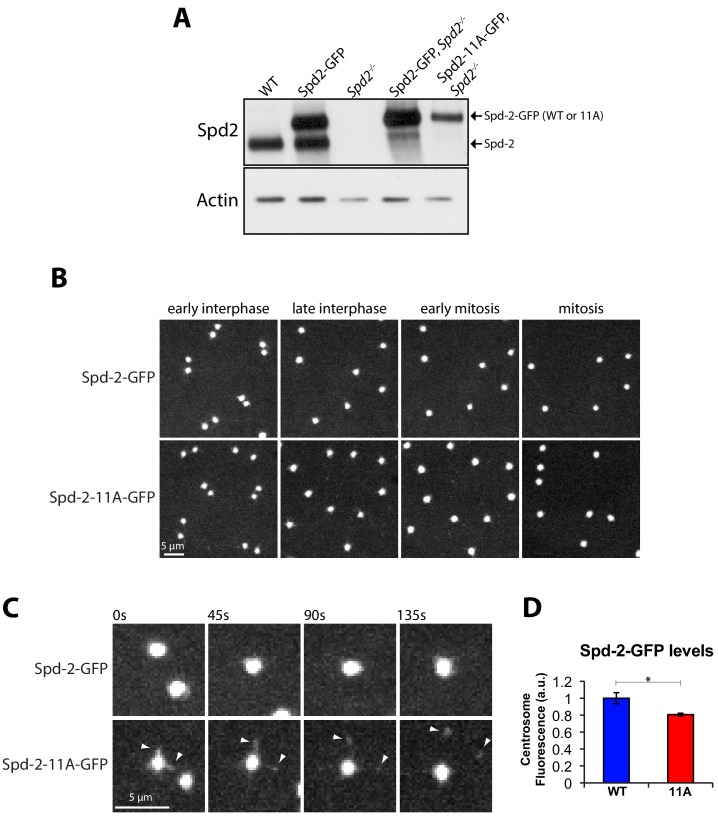
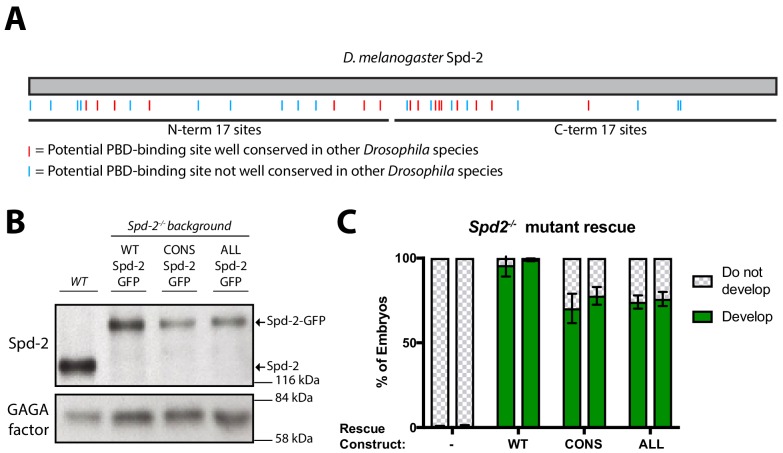
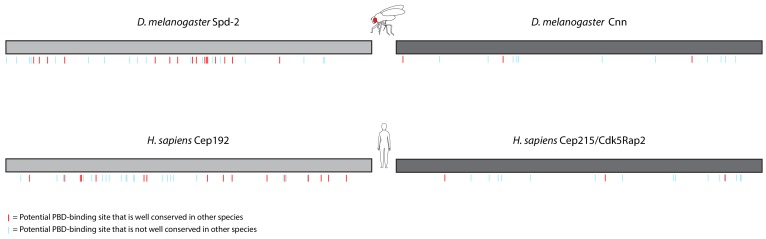
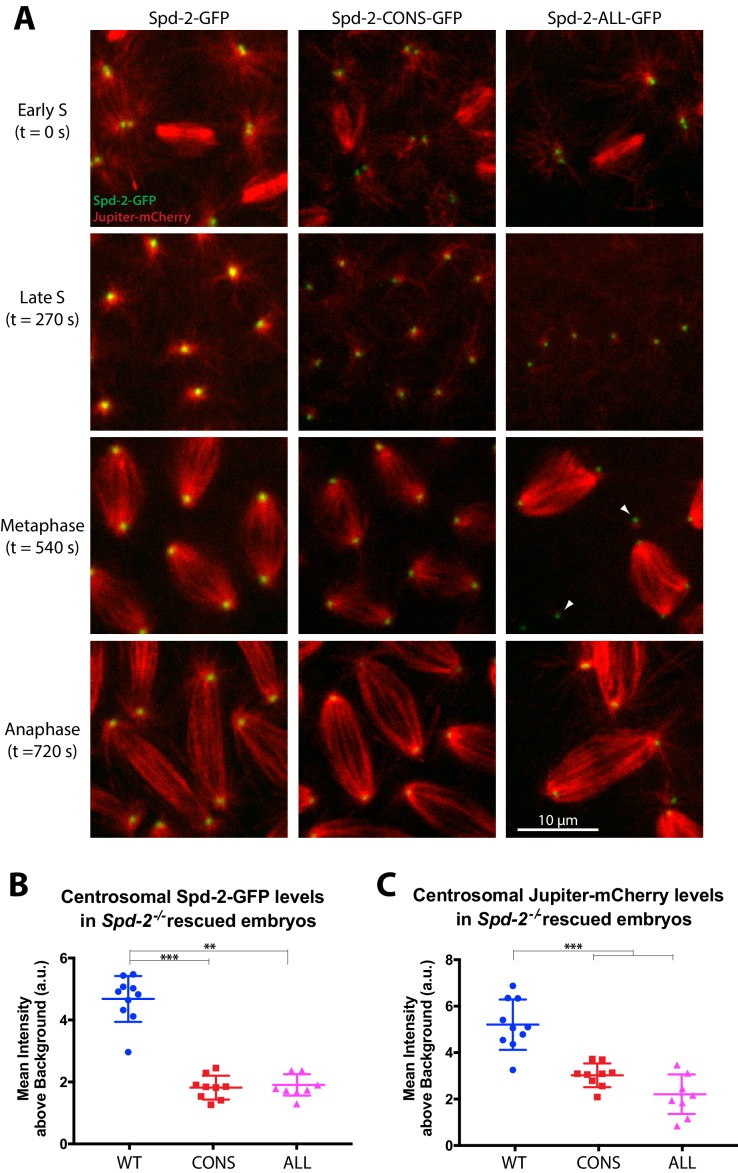
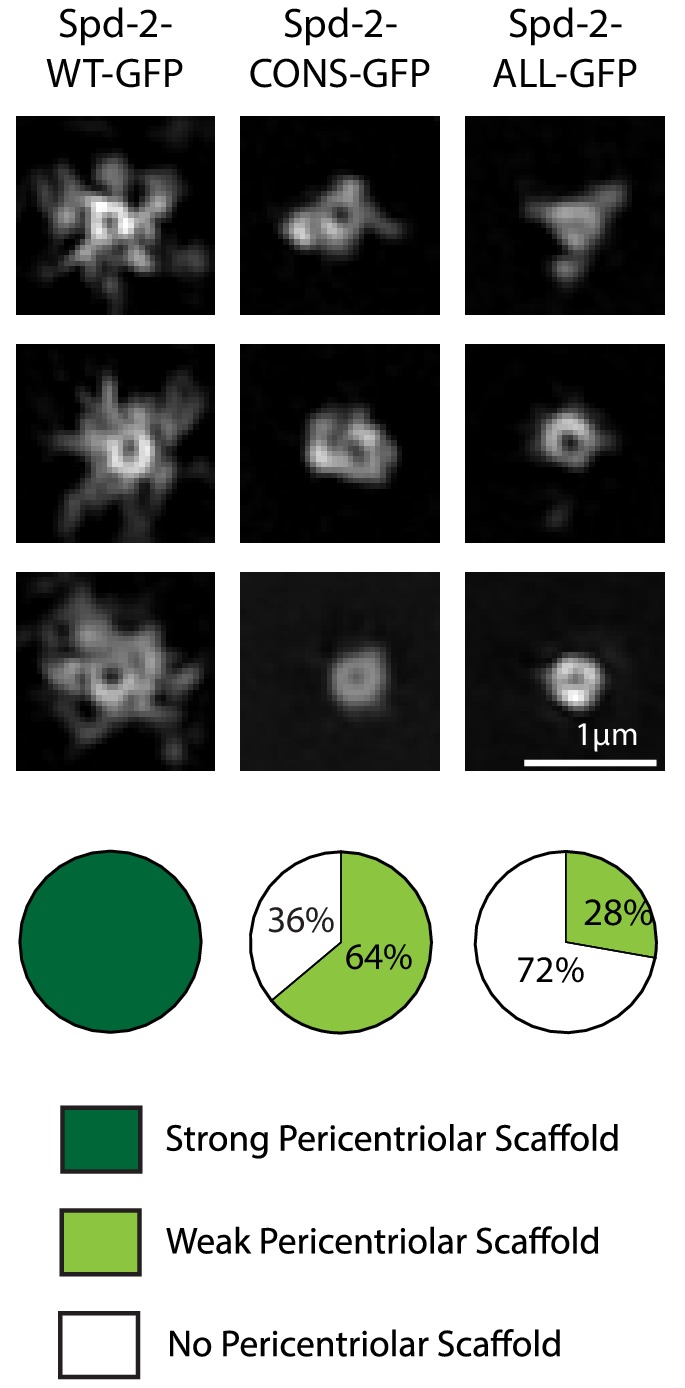
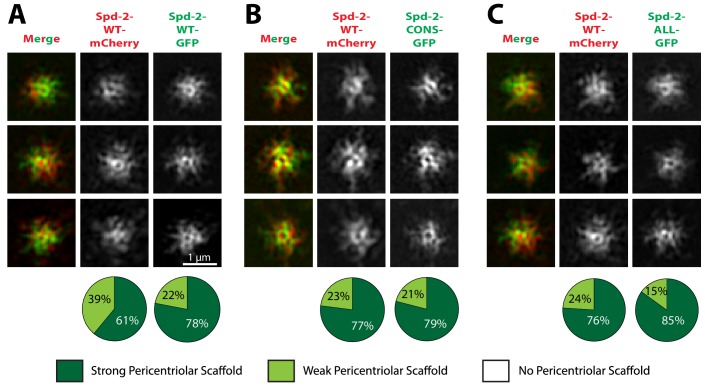
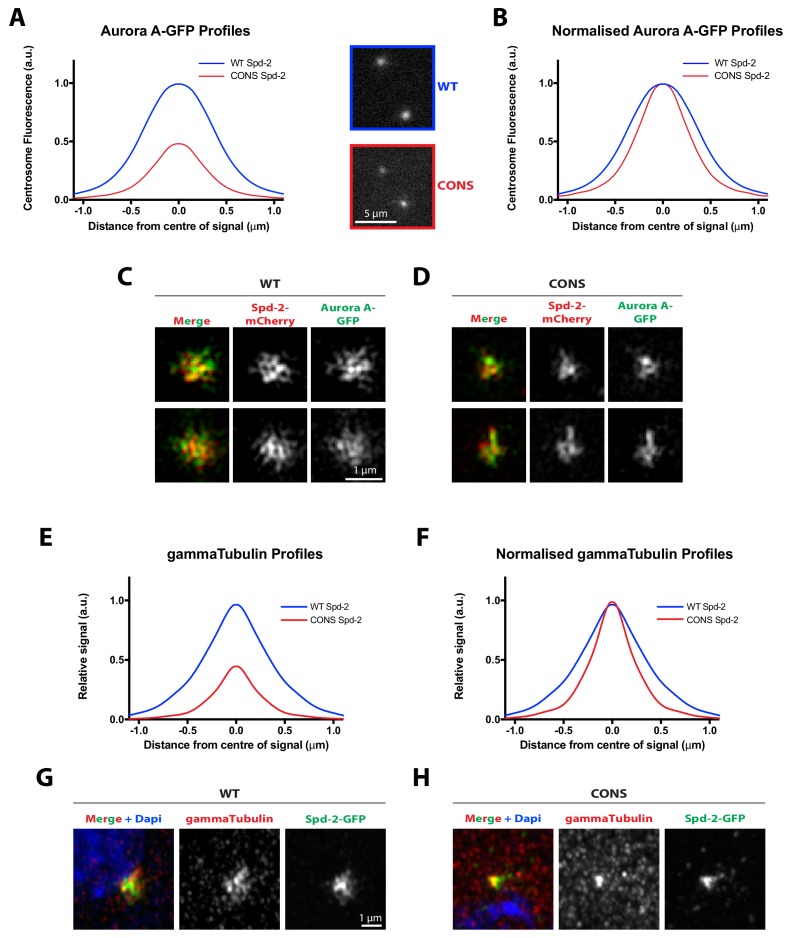
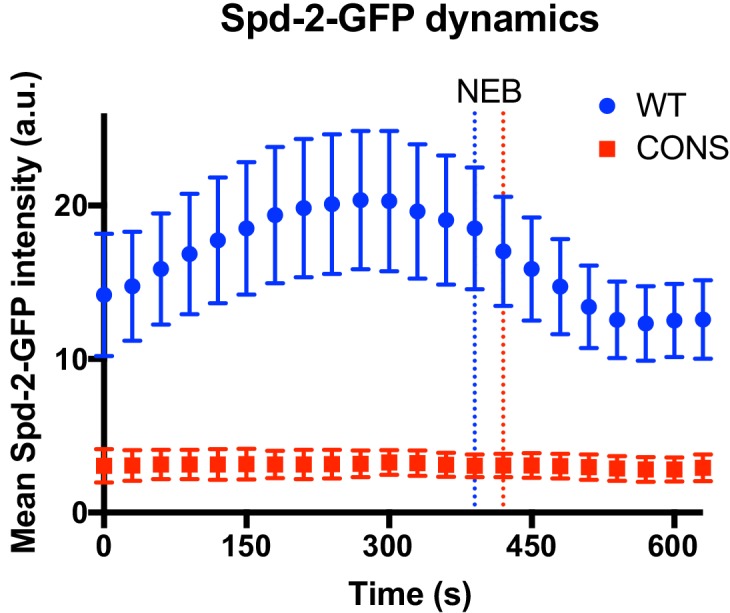
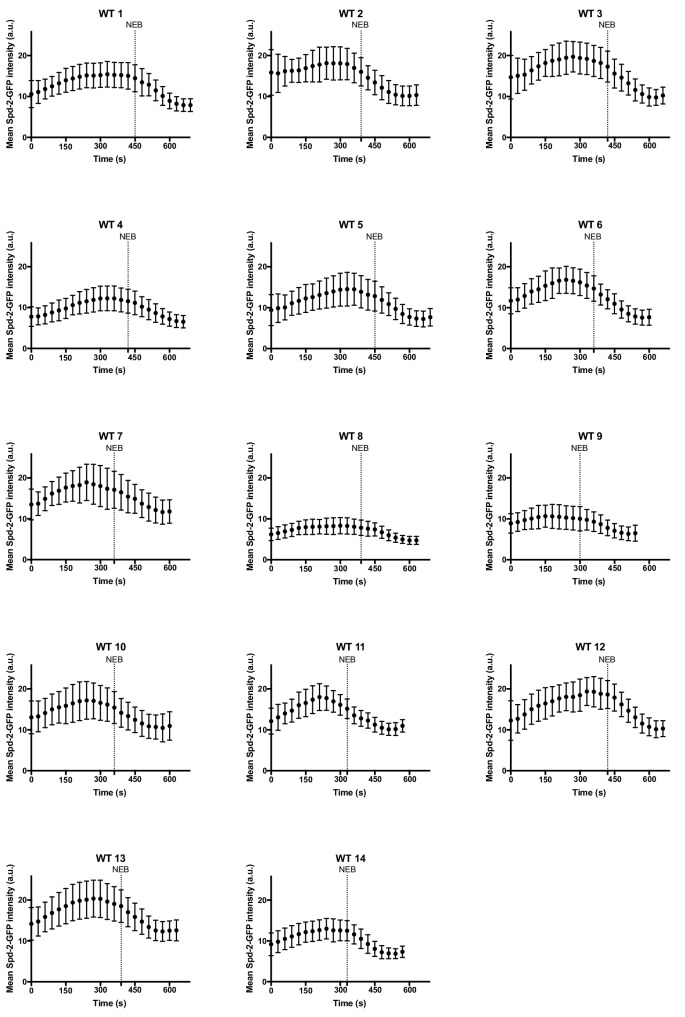
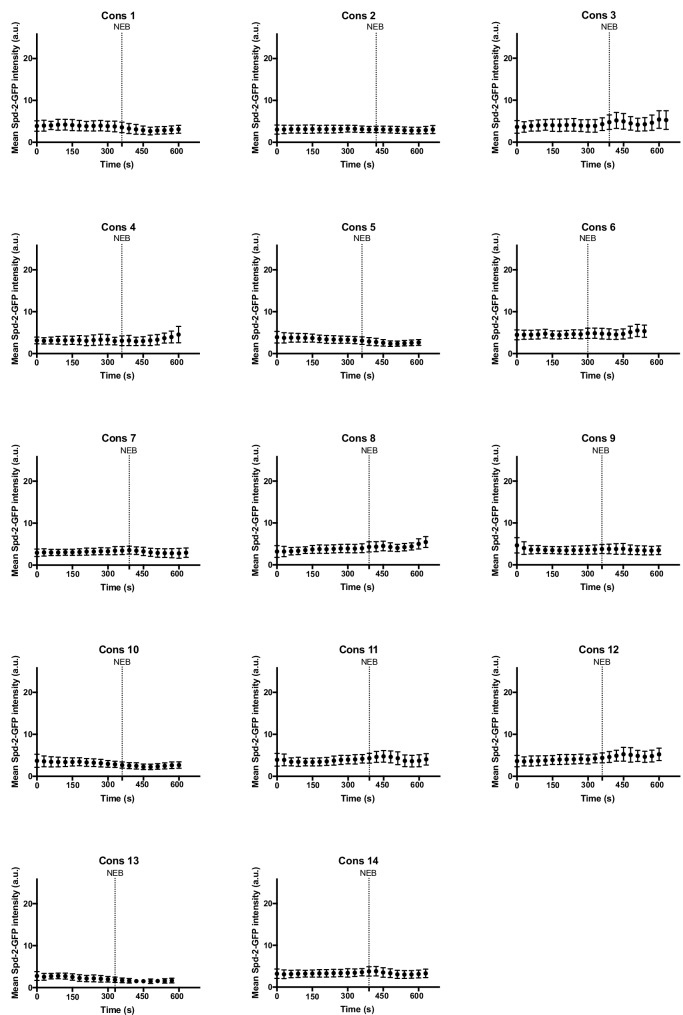
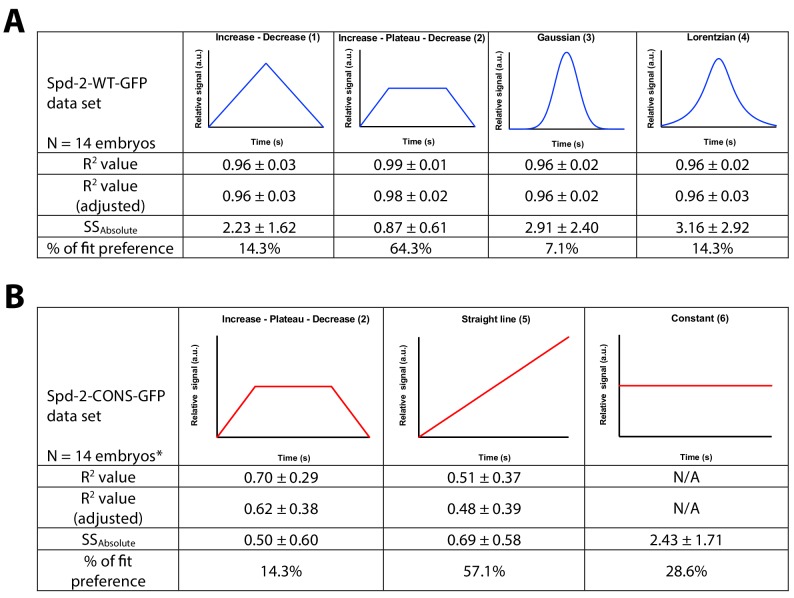
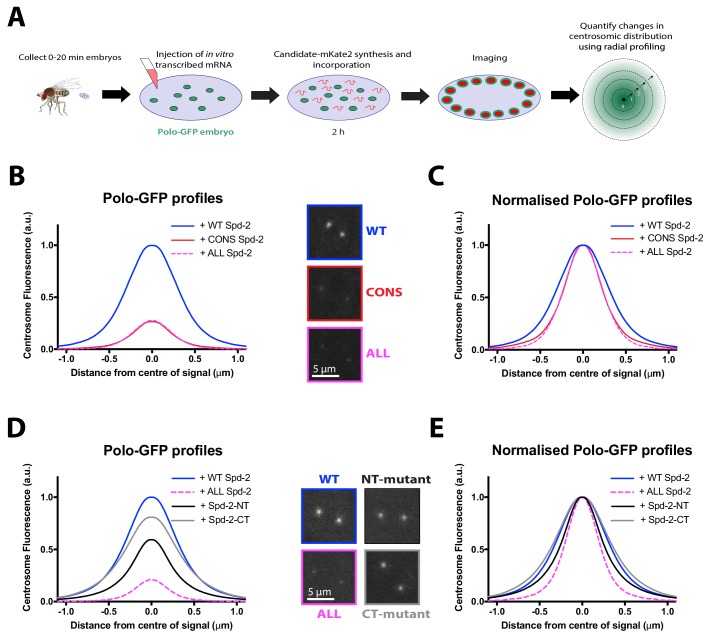
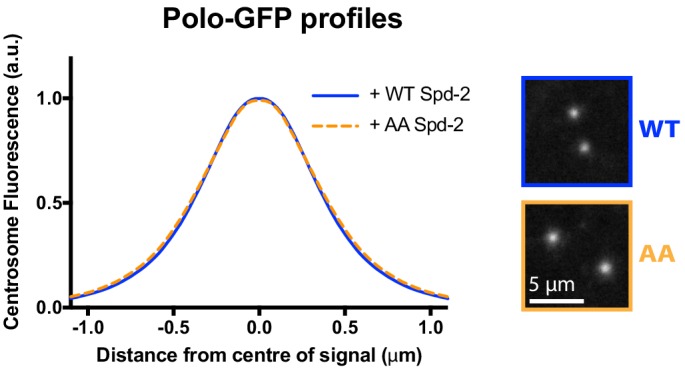
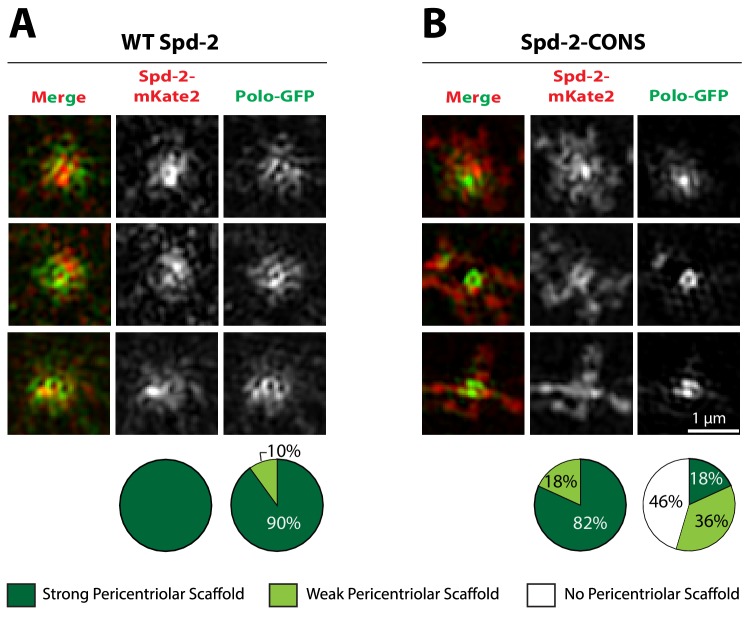
Centrosomes are formed when mother centrioles recruit pericentriolar material (PCM) around themselves. The PCM expands dramatically as cells prepare to enter mitosis (a process termed centrosome maturation), but it is unclear how this expansion is achieved. In flies, Spd-2 and Cnn are thought to form a scaffold around the mother centriole that recruits other components of the mitotic PCM, and the Polo-dependent phosphorylation of Cnn at the centrosome is crucial for scaffold assembly. Here, we show that, like Cnn, Spd-2 is specifically phosphorylated at centrosomes. This phosphorylation appears to create multiple phosphorylated S-S/T(p) motifs that allow Spd-2 to recruit Polo to the expanding scaffold. If the ability of Spd-2 to recruit Polo is impaired, the scaffold is initially assembled around the mother centriole, but it cannot expand outwards, and centrosome maturation fails. Our findings suggest that interactions between Spd-2, Polo and Cnn form a positive feedback loop that drives the dramatic expansion of the mitotic PCM in fly embryos.
中心体是在母中心粒周围招募中心粒周围物质 (PCM) 形成的。PCM 在细胞准备进入有丝分裂时会剧烈扩张(这个过程称为中心体成熟),但目前尚不清楚这种扩张是如何实现的。在果蝇中,Spd-2 和 Cnn 被认为在母中心粒周围形成一个支架,招募有丝分裂 PCM 的其他成分,而 Polo 依赖性的 Cnn 在中心体上的磷酸化对于支架组装至关重要。在这里,我们表明,像 Cnn 一样,Spd-2 也在中心体上被特异性磷酸化。这种磷酸化似乎产生了多个磷酸化的 S-S/T(p) 模体,使 Spd-2 能够招募 Polo 到扩展的支架上。如果 Spd-2 招募 Polo 的能力受损,支架最初会在母中心粒周围组装,但它不能向外扩展,中心体成熟失败。我们的发现表明,Spd-2、Polo 和 Cnn 之间的相互作用形成了一个正反馈回路,驱动了果蝇胚胎中有丝分裂 PCM 的剧烈扩张。