Department of Biotechnology and Life Science, Tokyo University of Agriculture and Technology, Tokyo, Japan.
JEM Utilization Center, Japan Aerospace Exploration Agency, Ibaraki, Japan.
Appl Environ Microbiol. 2019 Nov 27;85(24). doi: 10.1128/AEM.01692-19. Print 2019 Dec 15.
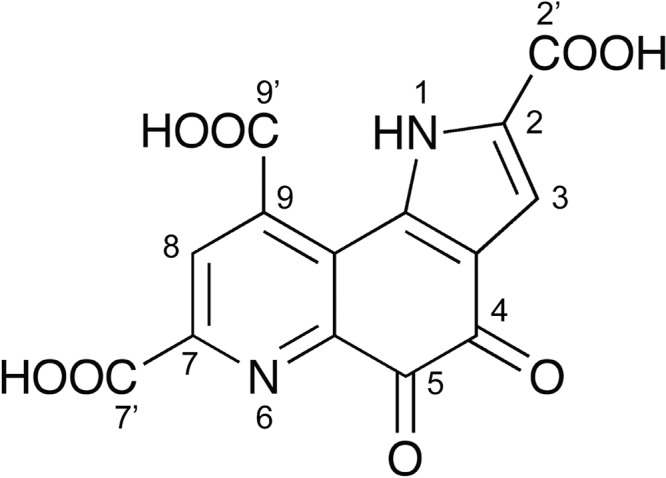
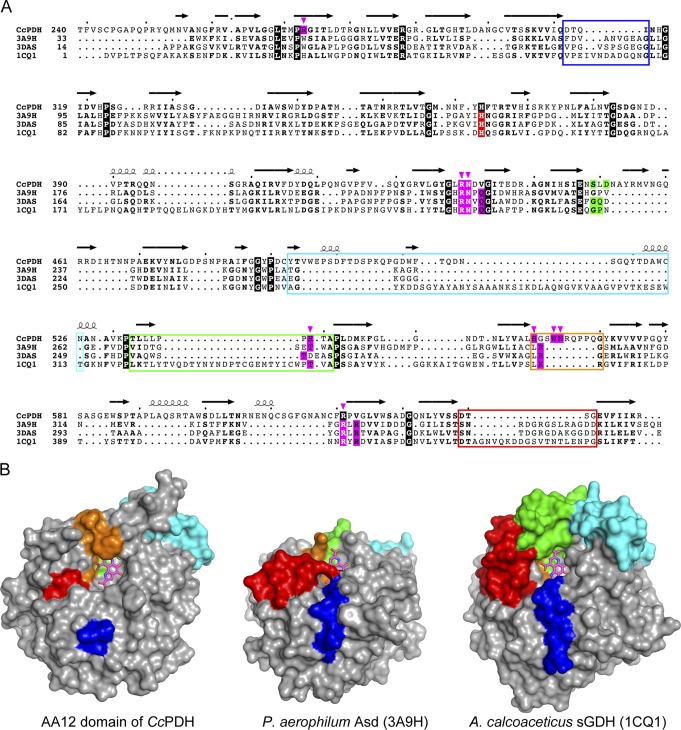
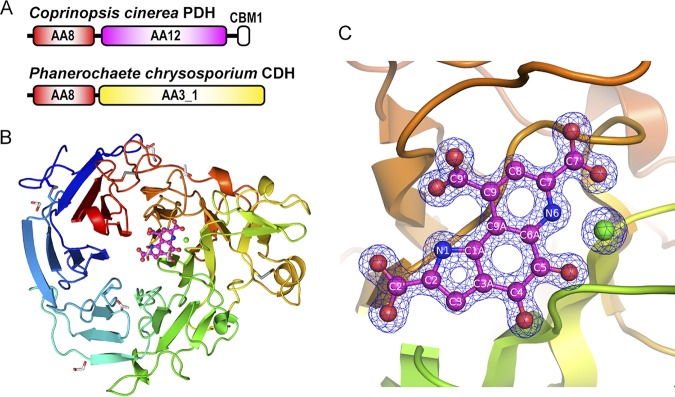
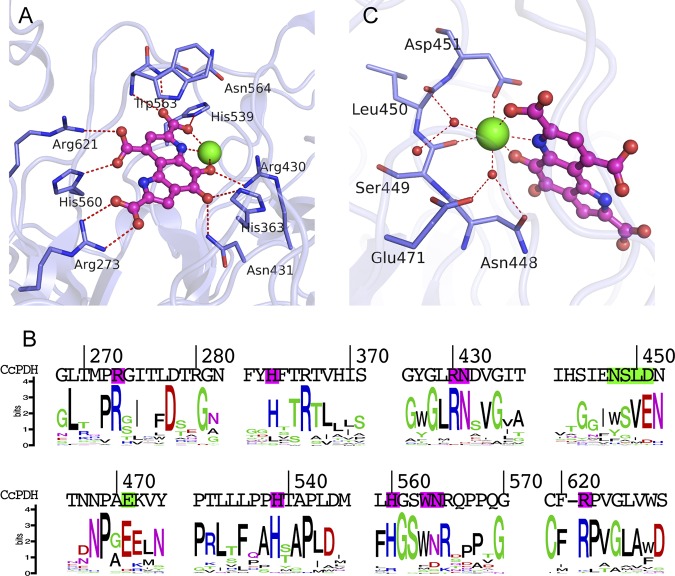
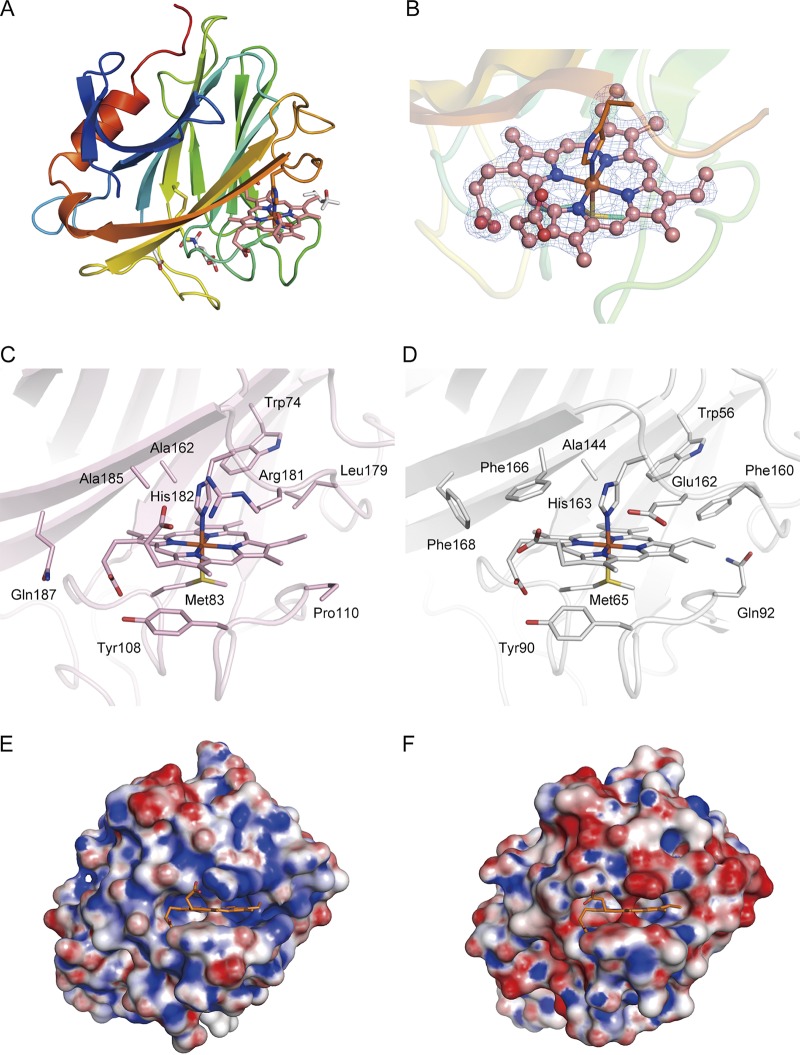
Pyrroloquinoline quinone (PQQ) was discovered as a redox cofactor of prokaryotic glucose dehydrogenases in the 1960s, and subsequent studies have demonstrated its importance not only in bacterial systems but also in higher organisms. We have previously reported a novel eukaryotic quinohemoprotein that exhibited PQQ-dependent catalytic activity in a eukaryote. The enzyme, pyranose dehydrogenase (PDH), from the filamentous fungus (PDH) of the Basidiomycete division, is composed of a catalytic PQQ-dependent domain classified as a member of the novel auxiliary activity family 12 (AA12), an AA8 cytochrome domain, and a family 1 carbohydrate-binding module (CBM1), as defined by the Carbohydrate-Active Enzymes (CAZy) database. Here, we present the crystal structures of the AA12 domain in its apo- and holo-forms and the AA8 domain of this enzyme. The crystal structures of the holo-AA12 domain bound to PQQ provide direct evidence that eukaryotes have PQQ-dependent enzymes. The AA12 domain exhibits a six-blade β-propeller fold that is also present in other known PQQ-dependent glucose dehydrogenases in bacteria. A loop structure around the active site and a calcium ion binding site are unique among the known structures of bacterial quinoproteins. The AA8 cytochrome domain has a positively charged area on its molecular surface, which is partly due to the propionate group of the heme interacting with Arg181; this feature differs from the characteristics of cytochrome in the AA8 domain of the fungal cellobiose dehydrogenase and suggests that this difference may affect the pH dependence of electron transfer. Pyrroloquinoline quinone (PQQ) is known as the "third coenzyme" following nicotinamide and flavin. PQQ-dependent enzymes have previously been found only in prokaryotes, and the existence of a eukaryotic PQQ-dependent enzyme was in doubt. In 2014, we found an enzyme in mushrooms that catalyzes the oxidation of various sugars in a PQQ-dependent manner and that was a PQQ-dependent enzyme found in eukaryotes. This paper presents the X-ray crystal structures of this eukaryotic PQQ-dependent quinohemoprotein, which show the active site, and identifies the amino acid residues involved in the binding of the cofactor PQQ. The presented X-ray structures reveal that the AA12 domain is in a binary complex with the coenzyme, clearly proving that PQQ-dependent enzymes exist in eukaryotes as well as prokaryotes. Because no biosynthetic system for PQQ has been reported in eukaryotes, future research on the symbiotic systems is expected.
吡咯并喹啉醌(PQQ)于 20 世纪 60 年代被发现为原核葡萄糖脱氢酶的氧化还原辅因子,随后的研究不仅证明了其在细菌系统中的重要性,也证明了其在高等生物中的重要性。我们之前报道了一种新型真核醌蛋白,它在真核生物中表现出依赖 PQQ 的催化活性。该酶来自担子菌门的丝状真菌,即戊糖氧化酶(PDH),由依赖 PQQ 的催化结构域、属于新型辅助活性家族 12(AA12)的结构域、AA8 细胞色素结构域和家族 1 碳水化合物结合模块(CBM1)组成,这是由碳水化合物活性酶(CAZy)数据库定义的。在这里,我们展示了该酶的 AA12 结构域在其apo 和 holo 形式以及 AA8 结构域的晶体结构。与 PQQ 结合的 holo-AA12 结构域的晶体结构提供了直接的证据,证明真核生物具有依赖 PQQ 的酶。AA12 结构域具有六叶β-桨叶折叠,也存在于细菌中其他已知的依赖 PQQ 的葡萄糖脱氢酶中。活性位点周围的环结构和钙离子结合位点在已知的细菌醌蛋白结构中是独特的。AA8 细胞色素结构域在其分子表面上具有带正电荷的区域,这部分归因于血红素的丙酸基团与 Arg181 相互作用;这一特征与真菌纤维二糖脱氢酶的 AA8 结构域中的细胞色素特征不同,并表明这种差异可能影响电子转移对 pH 的依赖性。吡咯并喹啉醌(PQQ)被称为继烟酰胺和黄素之后的“第三种辅酶”。以前只在原核生物中发现依赖 PQQ 的酶,而真核生物中是否存在依赖 PQQ 的酶还存在疑问。2014 年,我们在蘑菇中发现了一种酶,它以 PQQ 依赖的方式催化各种糖的氧化,这是一种在真核生物中发现的依赖 PQQ 的酶。本文介绍了这种真核 PQQ 依赖的醌蛋白的 X 射线晶体结构,显示了活性位点,并确定了与辅因子 PQQ 结合的氨基酸残基。所呈现的 X 射线结构表明,AA12 结构域与辅酶处于二元复合物中,这清楚地证明了依赖 PQQ 的酶不仅存在于原核生物中,也存在于真核生物中。由于在真核生物中没有报道 PQQ 的生物合成系统,因此预计未来将对共生系统进行研究。