The Sainsbury Laboratory, University of East Anglia, Norwich Research Park, Norwich, United Kingdom.
Department of Life Sciences, Imperial College London, London, United Kingdom.
Elife. 2019 Nov 27;8:e49956. doi: 10.7554/eLife.49956.
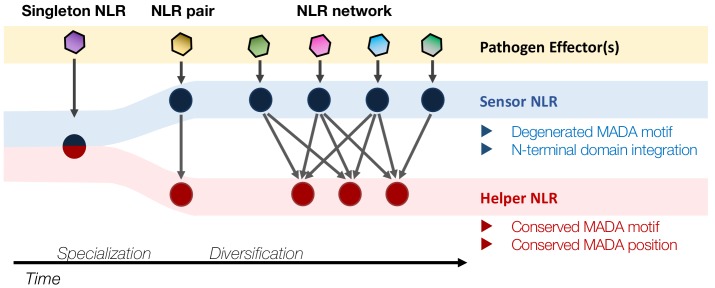
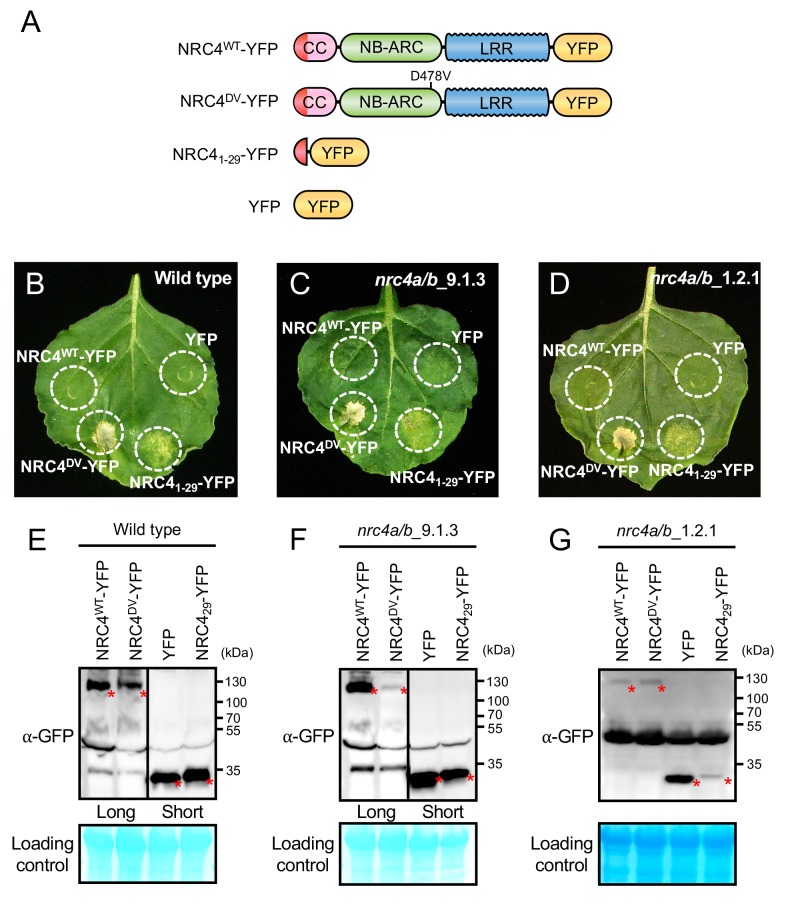
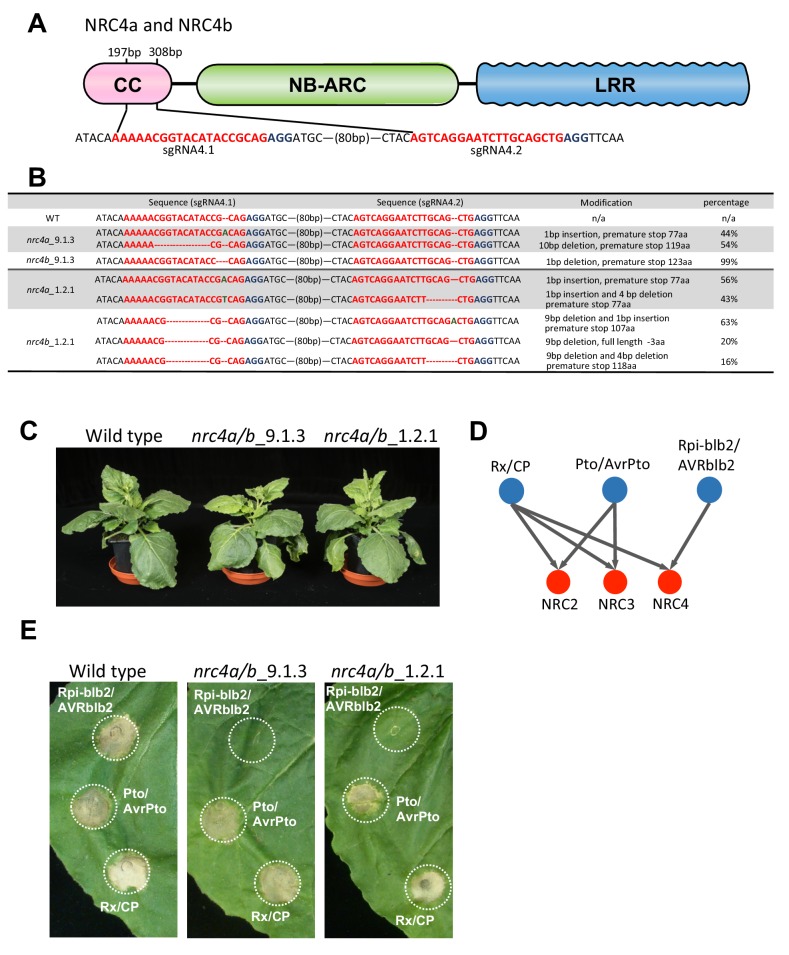
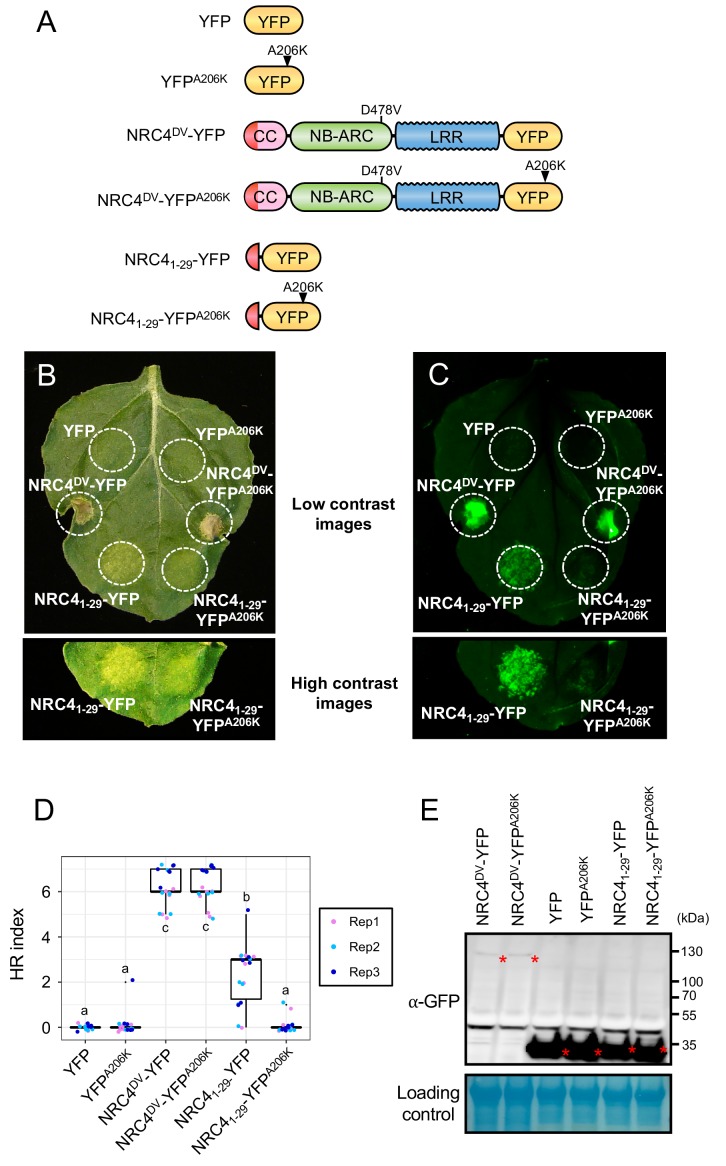
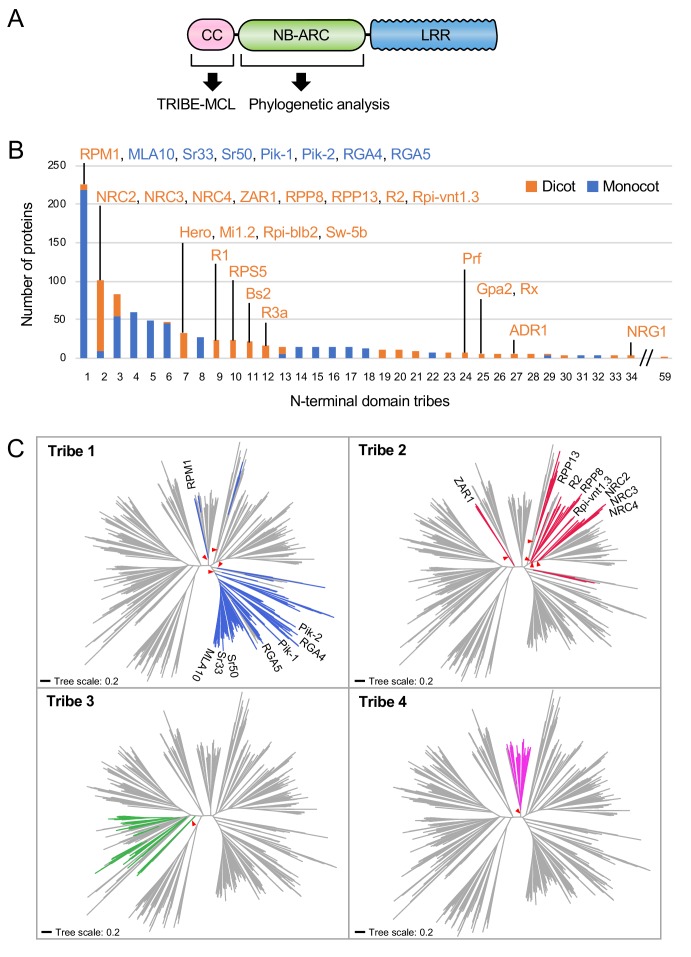
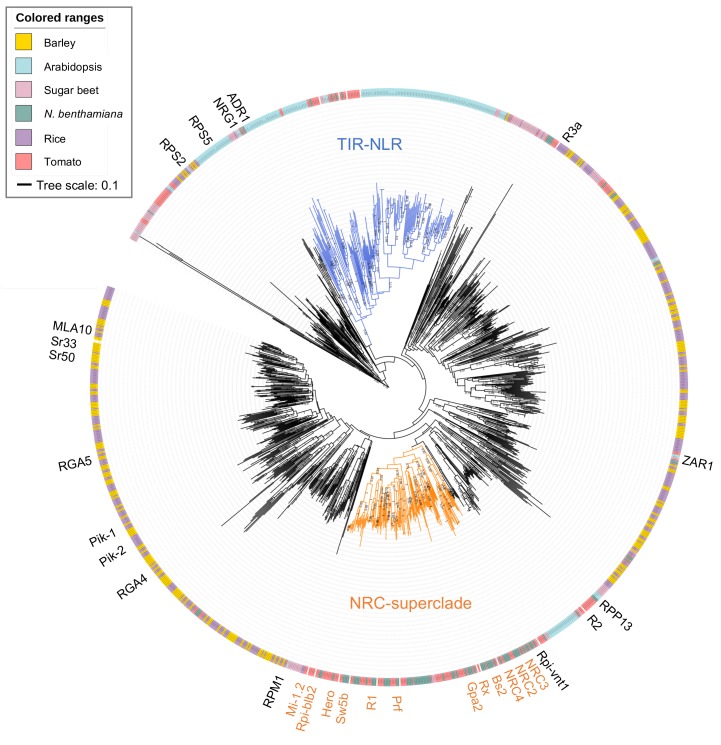
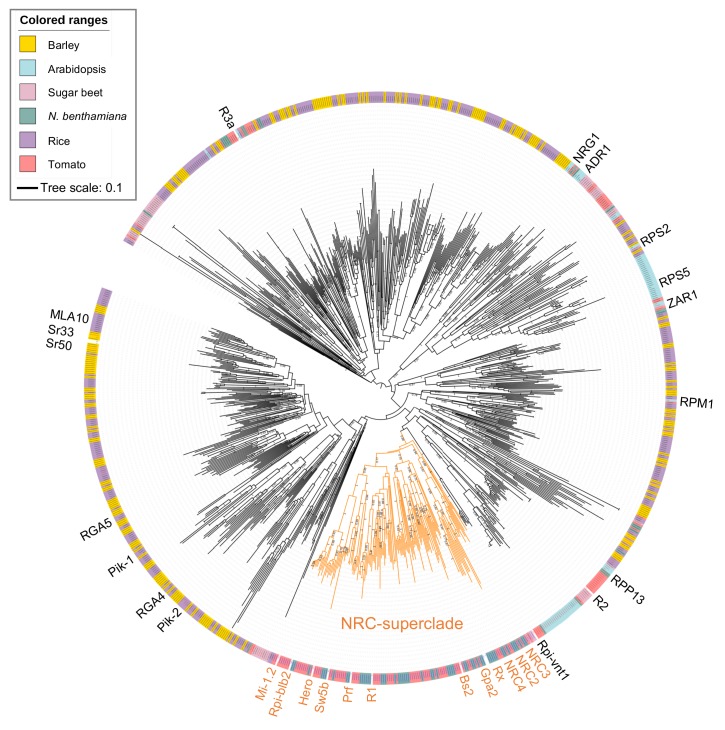
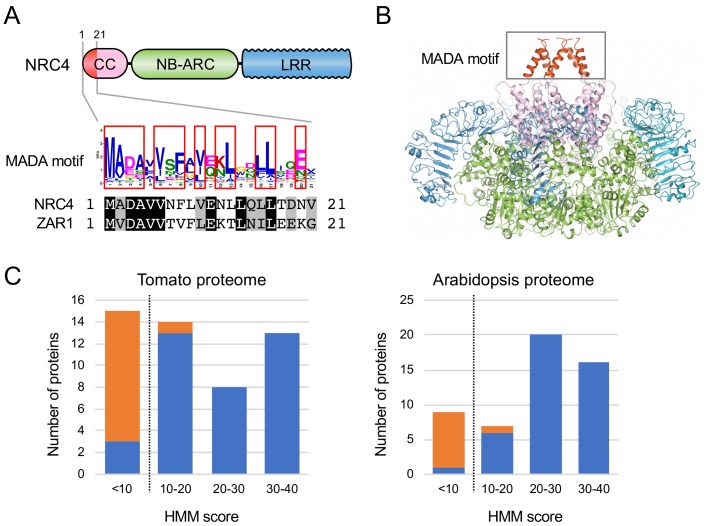
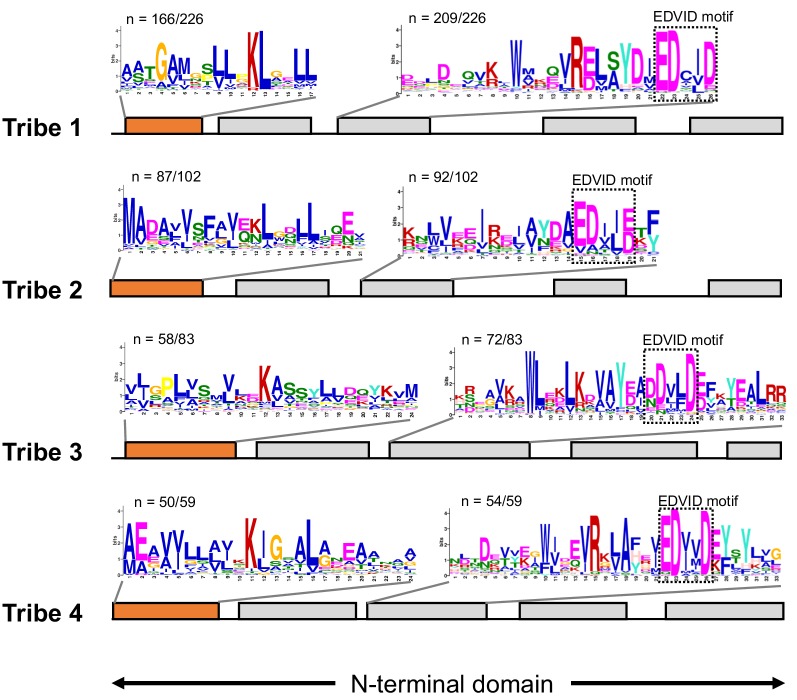
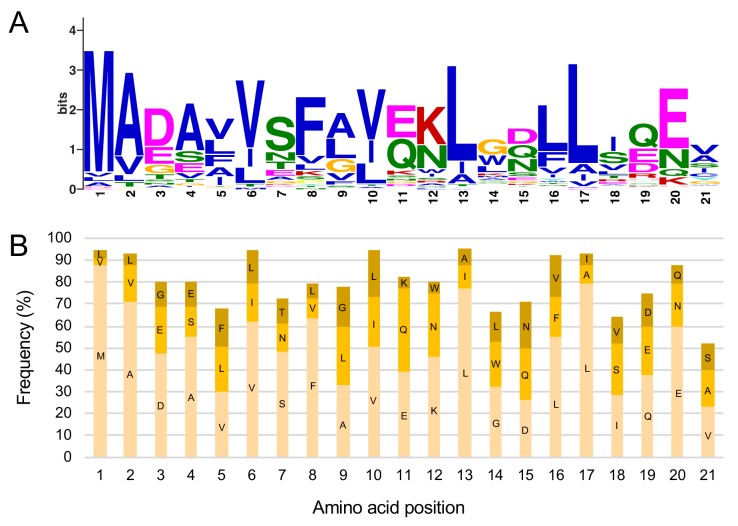
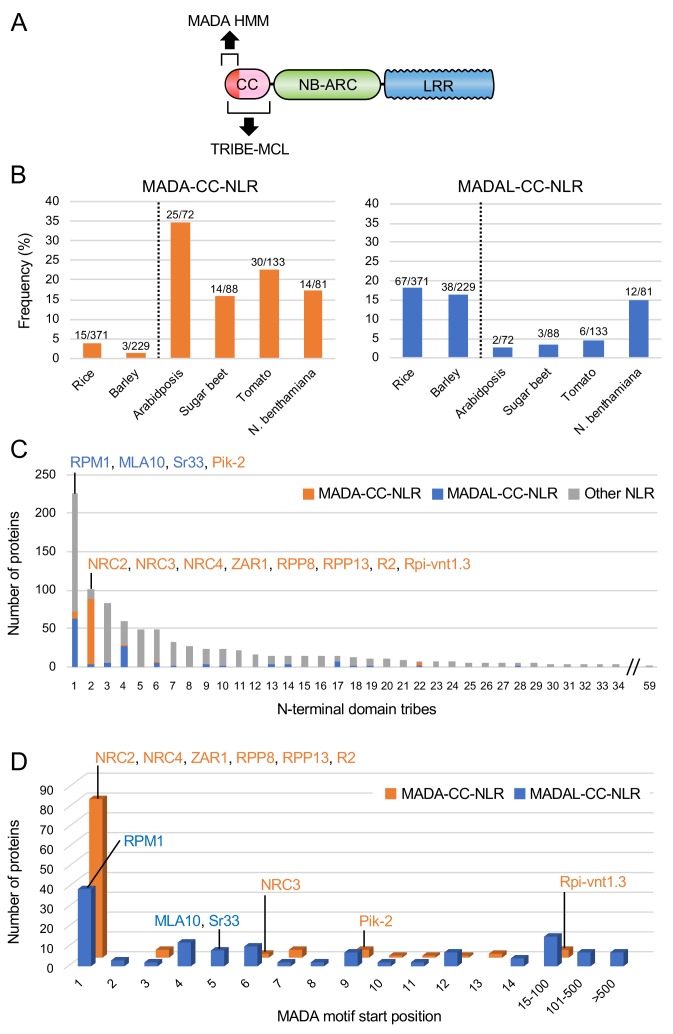
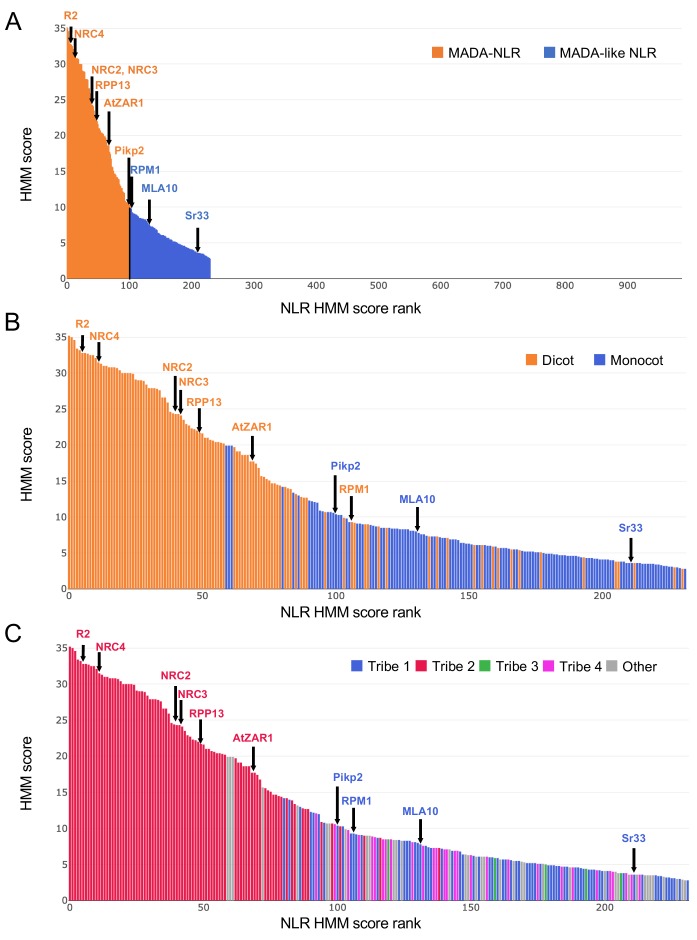
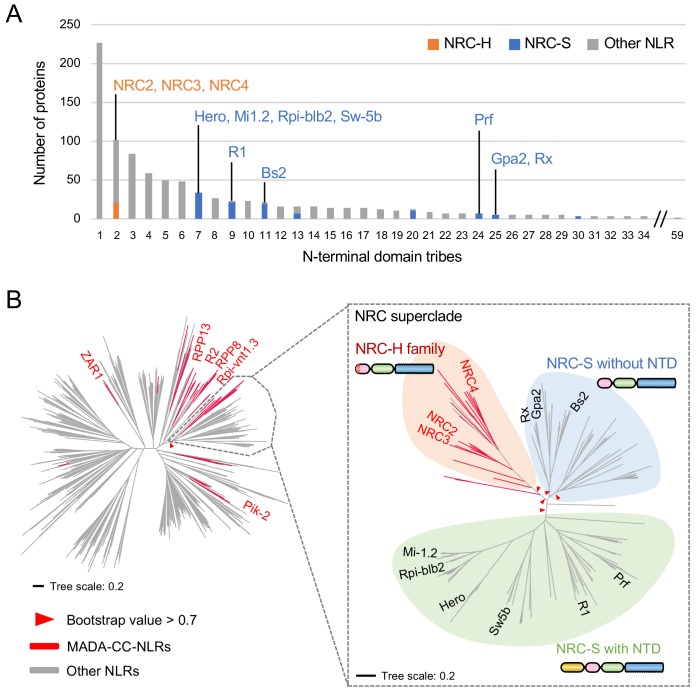
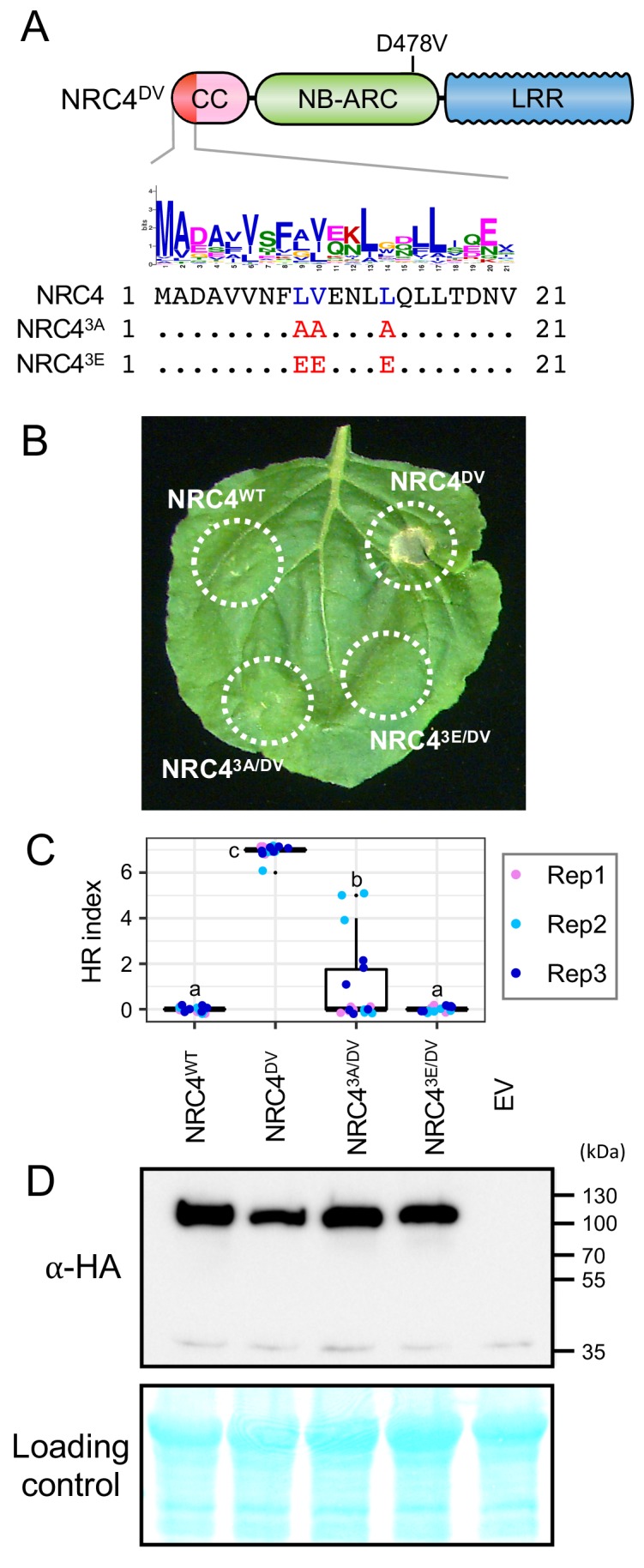
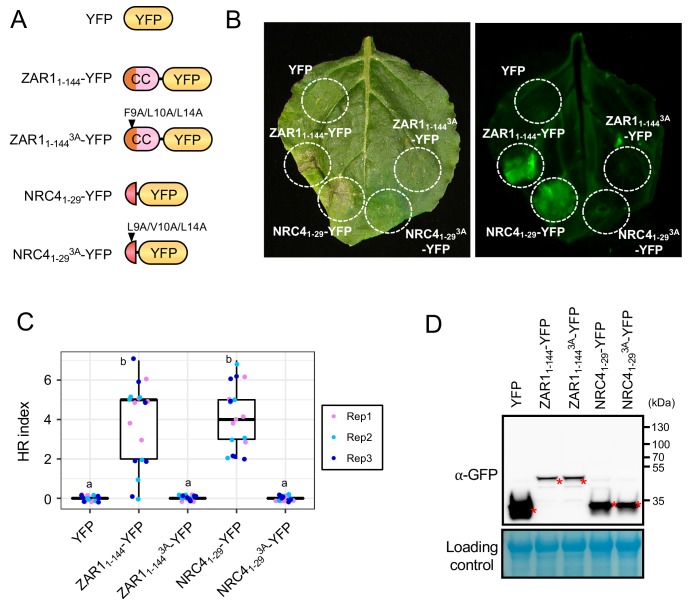
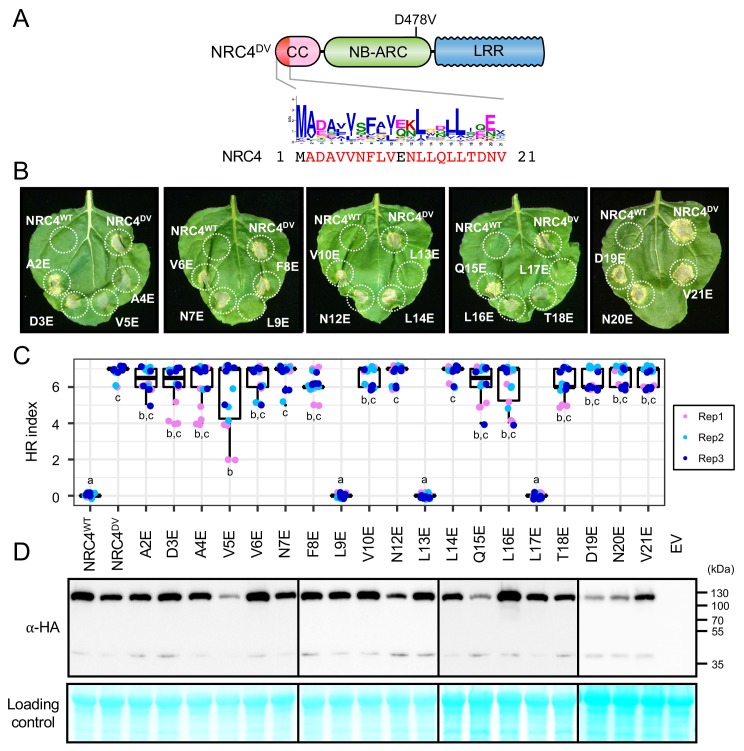
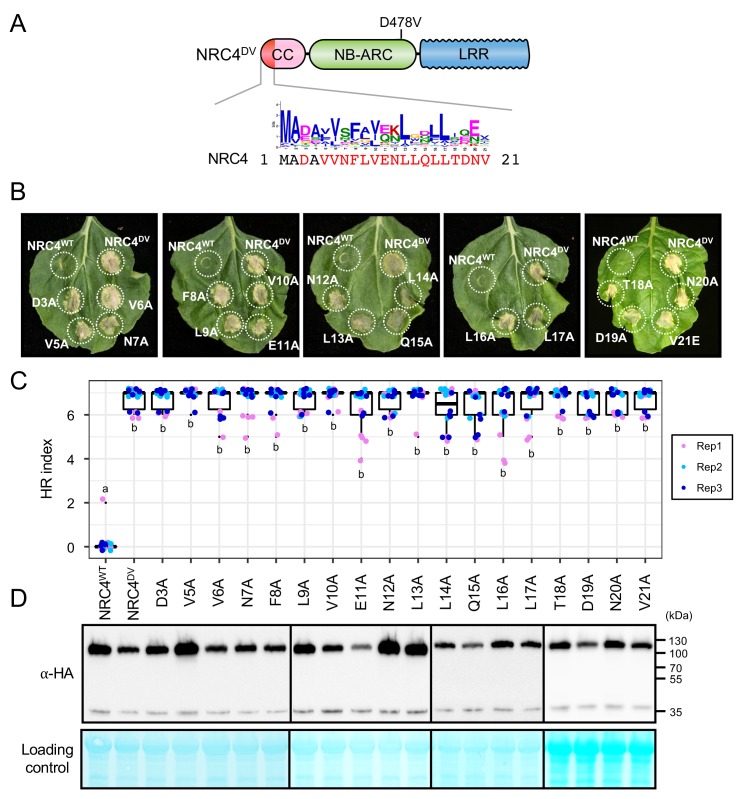
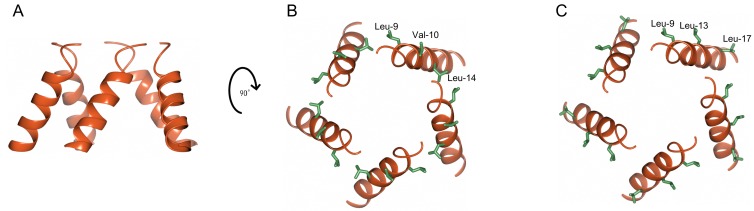
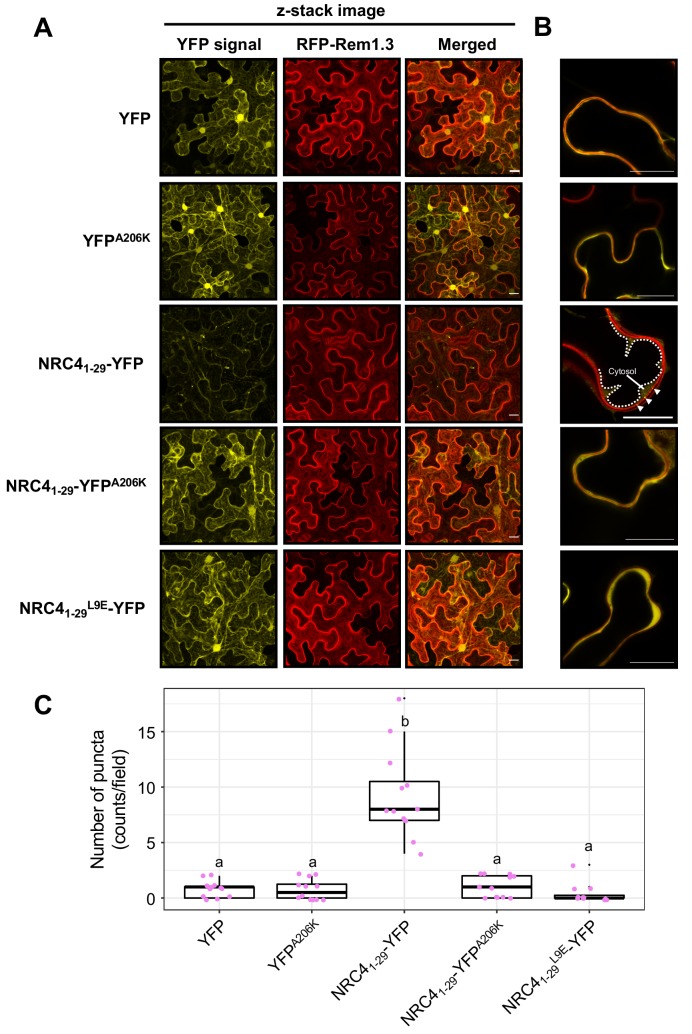
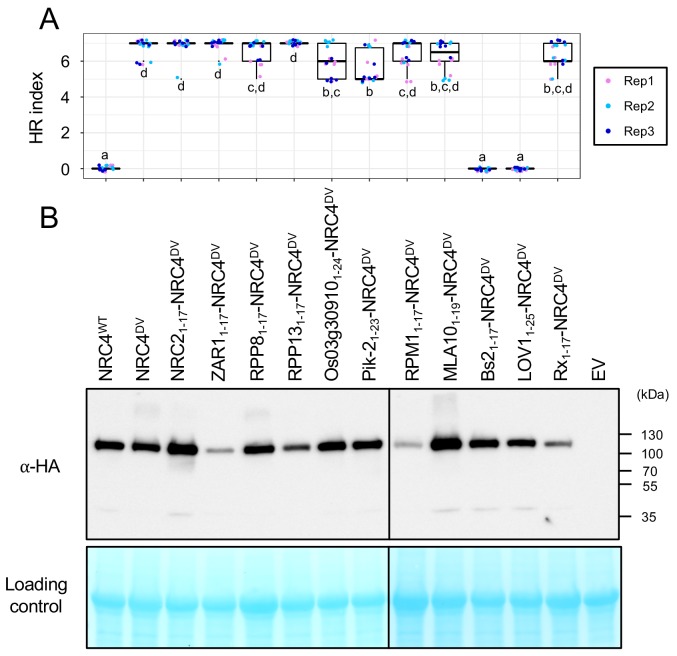
The molecular codes underpinning the functions of plant NLR immune receptors are poorly understood. We used in vitro Mu transposition to generate a random truncation library and identify the minimal functional region of NLRs. We applied this method to NRC4-a helper NLR that functions with multiple sensor NLRs within a Solanaceae receptor network. This revealed that the NRC4 N-terminal 29 amino acids are sufficient to induce hypersensitive cell death. This region is defined by the consensus MADAxVSFxVxKLxxLLxxEx (MADA motif) that is conserved at the N-termini of NRC family proteins and ~20% of coiled-coil (CC)-type plant NLRs. The MADA motif matches the N-terminal α1 helix of Arabidopsis NLR protein ZAR1, which undergoes a conformational switch during resistosome activation. Immunoassays revealed that the MADA motif is functionally conserved across NLRs from distantly related plant species. NRC-dependent sensor NLRs lack MADA sequences indicating that this motif has degenerated in sensor NLRs over evolutionary time.
植物 NLR 免疫受体功能的分子密码知之甚少。我们使用体外 Mu 转座生成随机截断文库,并鉴定 NLR 的最小功能区域。我们将这种方法应用于 NRC4——一种与茄科受体网络中的多个传感器 NLR 一起发挥作用的辅助 NLR。这表明 NRC4 的 N 端 29 个氨基酸足以诱导过敏细胞死亡。该区域由保守的 NRC 家族蛋白和~20%的卷曲螺旋 (CC)-型植物 NLR 的 N 末端的共有 MADAxVSFxVxKLxxLLxxEx (MADA 基序) 定义。MADA 基序与拟南芥 NLR 蛋白 ZAR1 的 N 端α1 螺旋匹配,该螺旋在抵抗体激活过程中发生构象转变。免疫测定显示,MADA 基序在来自不同植物物种的 NLR 中具有功能保守性。依赖 NRC 的传感器 NLR 缺乏 MADA 序列,表明该基序在进化过程中已经在传感器 NLR 中退化。