Institute of Medical Radiation Biology, University of Duisburg-Essen Medical School, Essen, Germany.
Nucleic Acids Res. 2020 Feb 28;48(4):1905-1924. doi: 10.1093/nar/gkz1167.
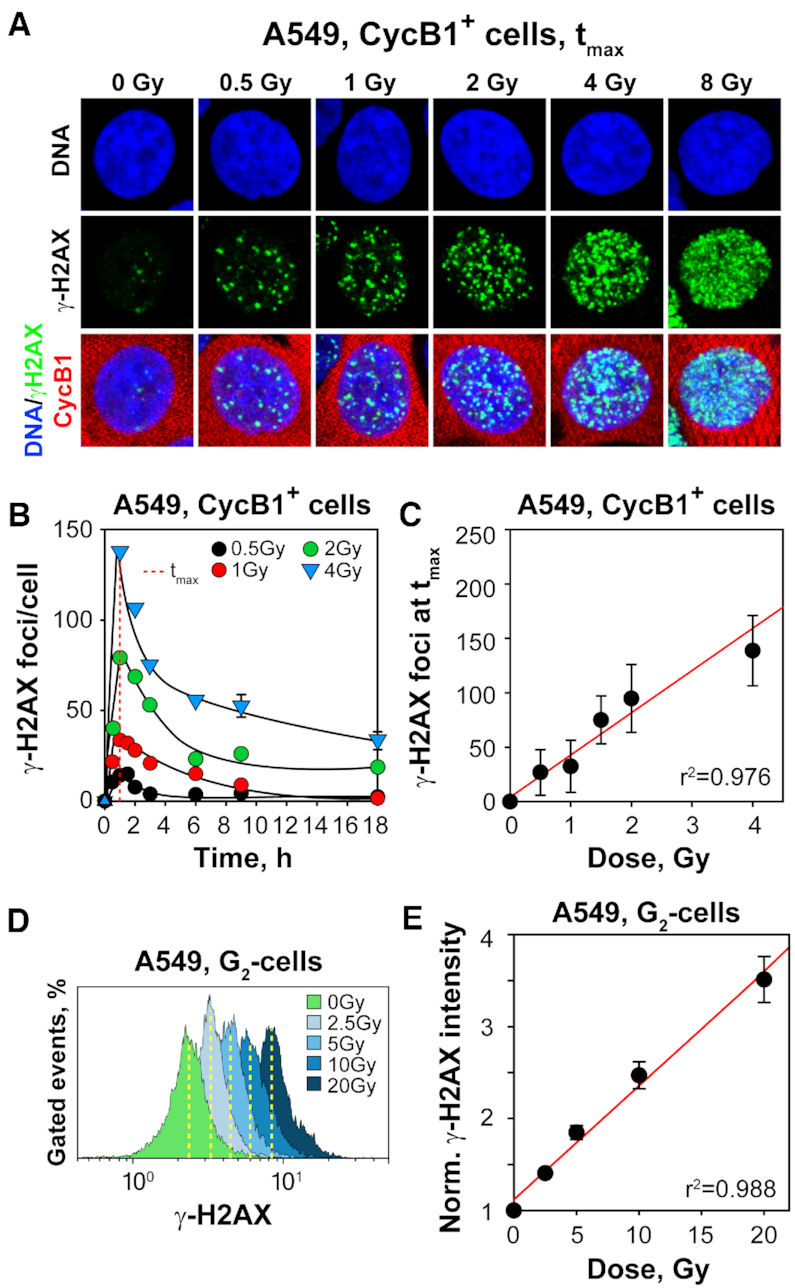
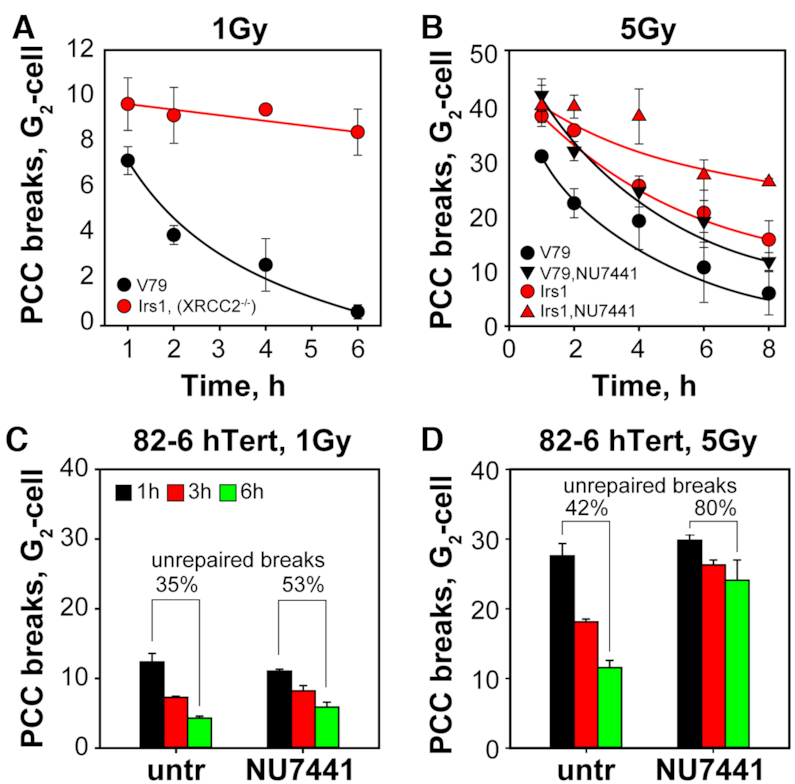
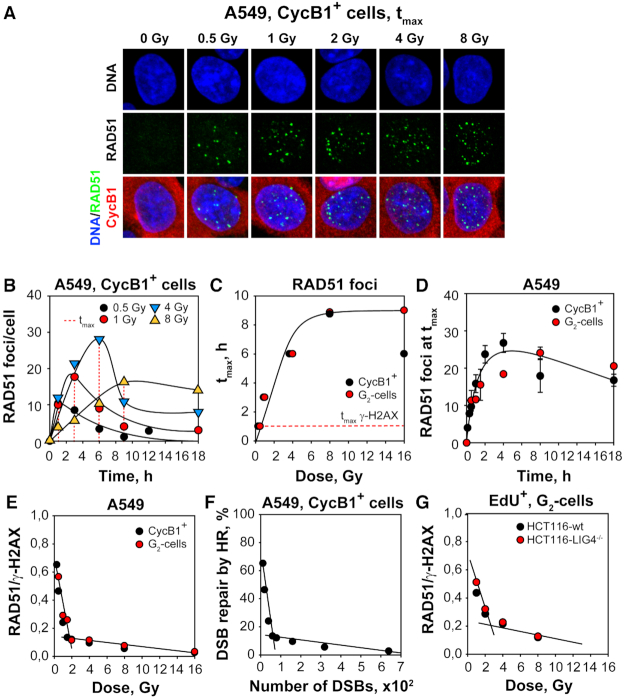
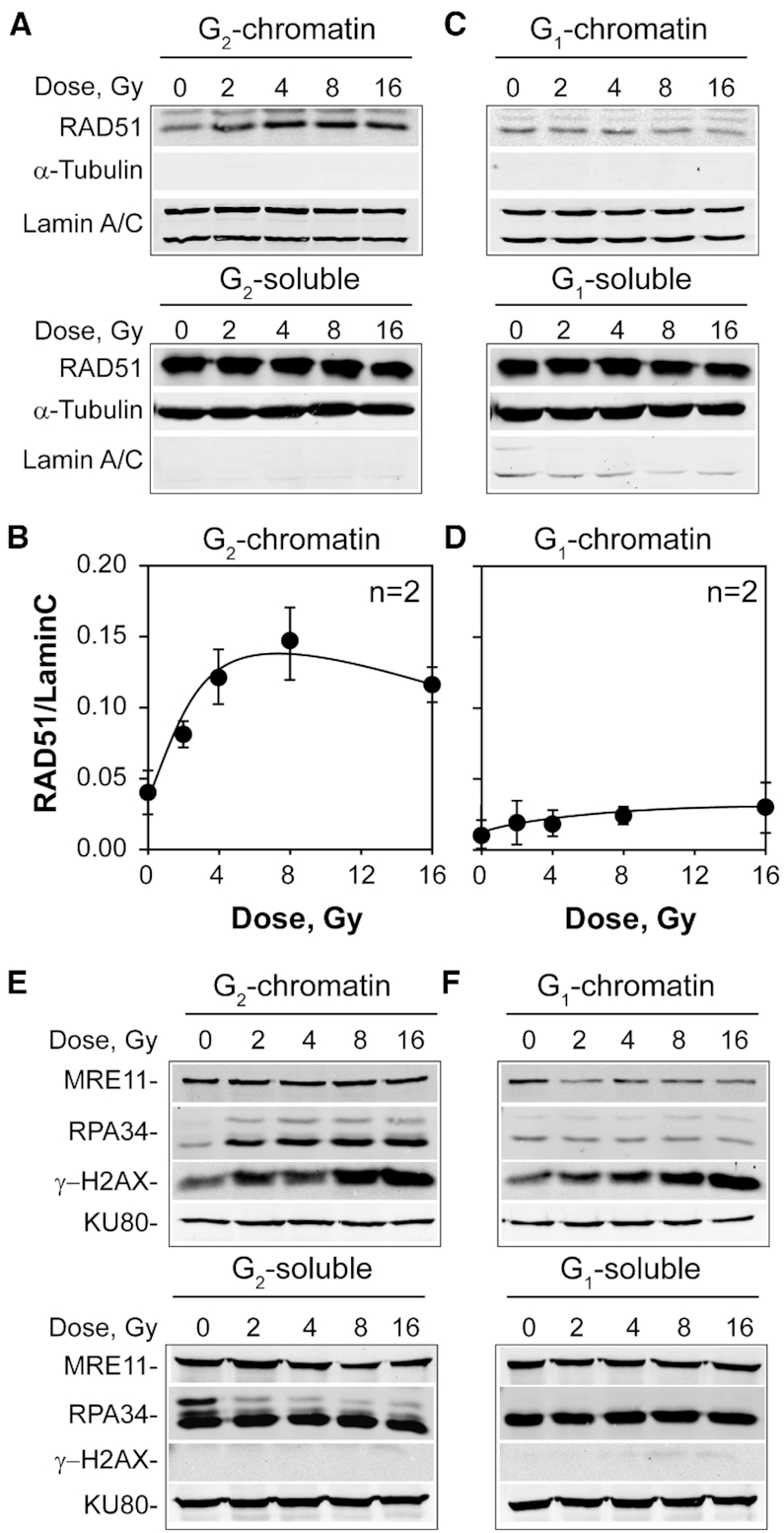
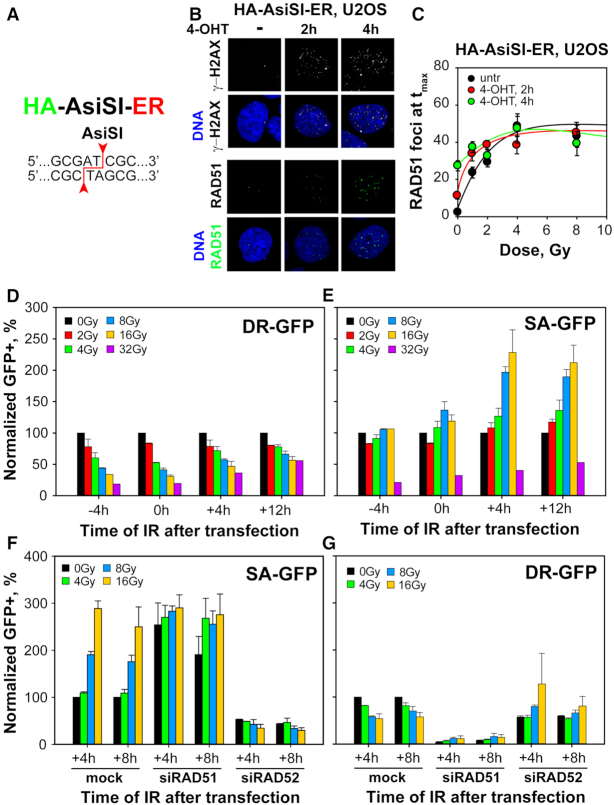
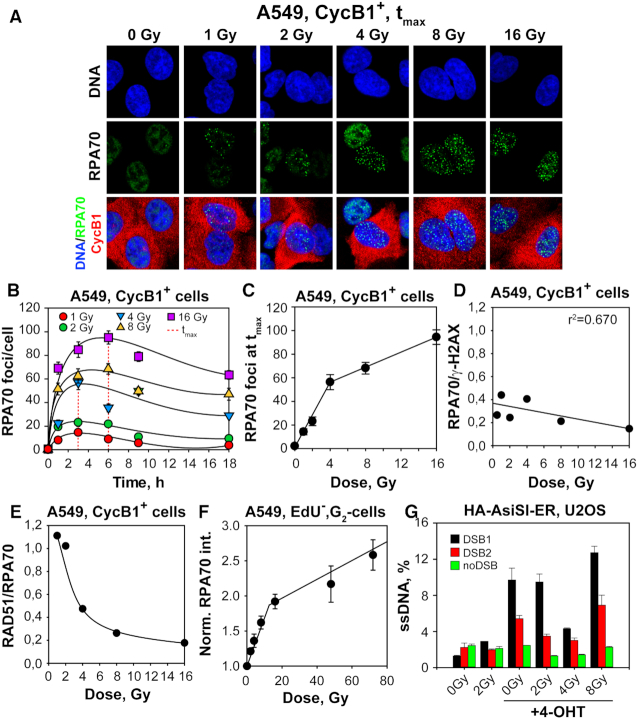
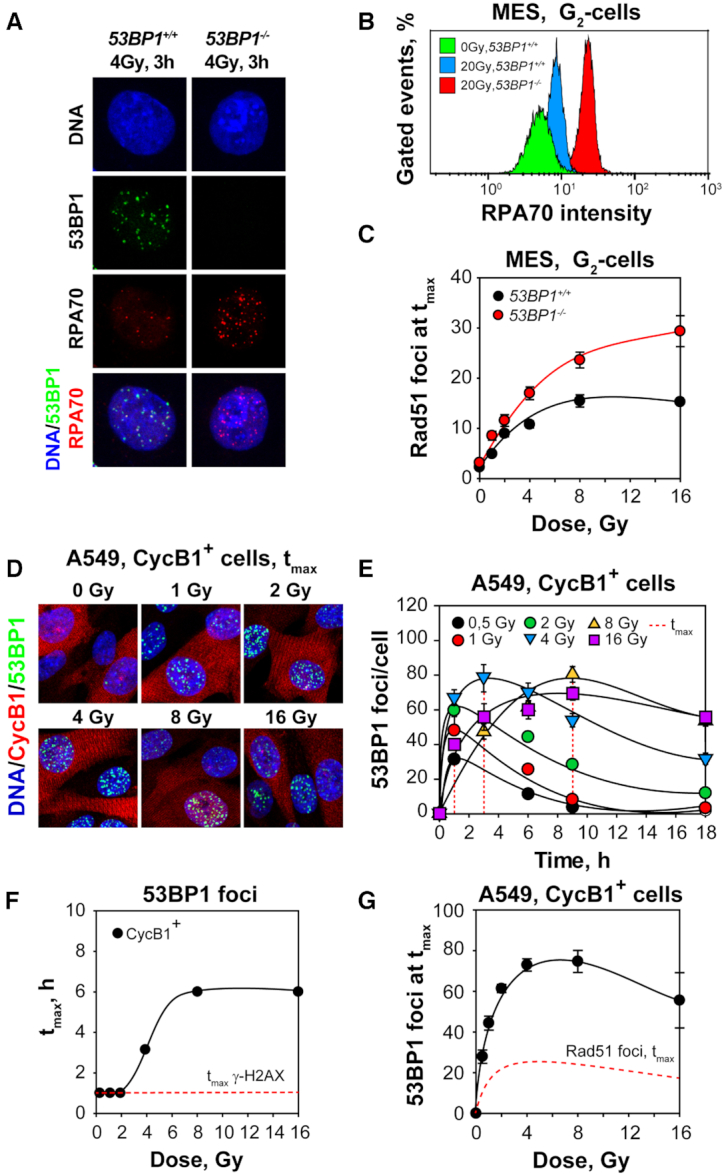
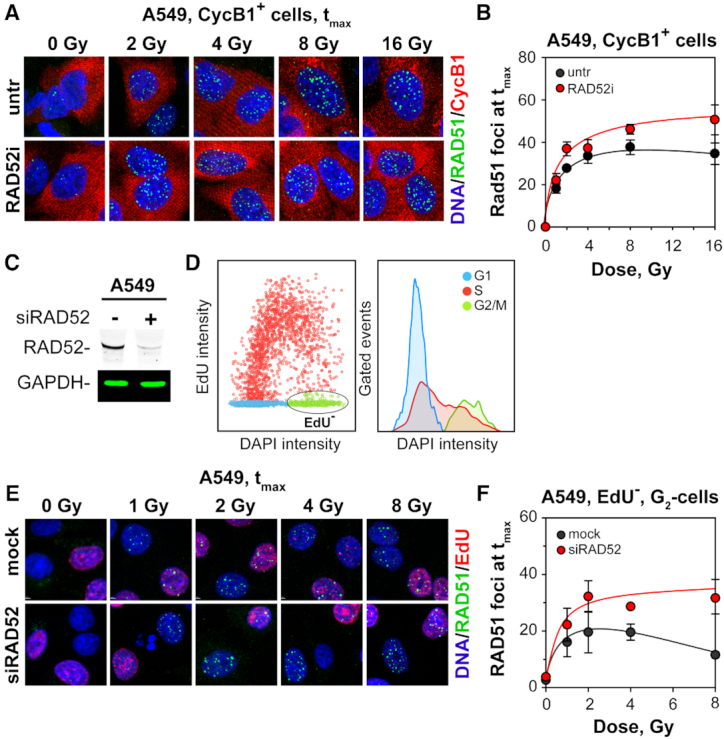
In vertebrates, genomic DNA double-strand breaks (DSBs) are removed by non-homologous end-joining processes: classical non-homologous end-joining (c-NHEJ) and alternative end-joining (alt-EJ); or by homology-dependent processes: gene-conversion (GC) and single-strand annealing (SSA). Surprisingly, these repair pathways are not real alternative options restoring genome integrity with equal efficiency, but show instead striking differences in speed, accuracy and cell-cycle-phase dependence. As a consequence, engagement of one pathway may be associated with processing-risks for the genome absent from another pathway. Characterization of engagement-parameters and their consequences is, therefore, essential for understanding effects on the genome of DSB-inducing agents, such as ionizing-radiation (IR). Here, by addressing pathway selection in G2-phase, we discover regulatory confinements in GC with consequences for SSA- and c-NHEJ-engagement. We show pronounced suppression of GC with increasing DSB-load that is not due to RAD51 availability and which is delimited but not defined by 53BP1 and RAD52. Strikingly, at low DSB-loads, GC repairs ∼50% of DSBs, whereas at high DSB-loads its contribution is undetectable. Notably, with increasing DSB-load and the associated suppression of GC, SSA gains ground, while alt-EJ is suppressed. These observations explain earlier, apparently contradictory results and advance our understanding of logic and mechanisms underpinning the wiring between DSB repair pathways.
在脊椎动物中,基因组 DNA 双链断裂 (DSBs) 通过非同源末端连接过程被修复:经典的非同源末端连接 (c-NHEJ) 和替代末端连接 (alt-EJ);或通过同源依赖过程:基因转换 (GC) 和单链退火 (SSA)。令人惊讶的是,这些修复途径并不是真正的替代选择,它们以相同的效率恢复基因组完整性,而是在速度、准确性和细胞周期依赖性方面表现出显著差异。因此,一种途径的参与可能与另一种途径不存在的基因组处理风险有关。因此,对参与参数及其后果的特征描述对于理解 DSB 诱导剂(如电离辐射 (IR))对基因组的影响至关重要。在这里,通过在 G2 期解决途径选择问题,我们发现 GC 中的调控限制,对 SSA 和 c-NHEJ 的参与具有后果。我们发现,随着 DSB 负荷的增加,GC 的抑制作用明显,这不是由于 RAD51 的可用性引起的,而是受到 53BP1 和 RAD52 的限制而不是定义。引人注目的是,在低 DSB 负荷下,GC 修复了约 50%的 DSB,而在高 DSB 负荷下,其贡献则无法检测到。值得注意的是,随着 DSB 负荷的增加和 GC 的抑制,SSA 占据了优势,而 alt-EJ 则被抑制。这些观察结果解释了早期似乎相互矛盾的结果,并提高了我们对 DSB 修复途径之间逻辑和机制的理解。