Université de Paris, INSERM U1016, UMR 8104 CNRS, Institut Cochin, Equipe Labellisée Ligue Contre le Cancer, France.
CNRS UMR 8200, Gustave-Roussy, Université Paris-Saclay, 114 rue Edouard Vaillant, 94805 Villejuif, France.
Nucleic Acids Res. 2022 Mar 21;50(5):2651-2666. doi: 10.1093/nar/gkac073.
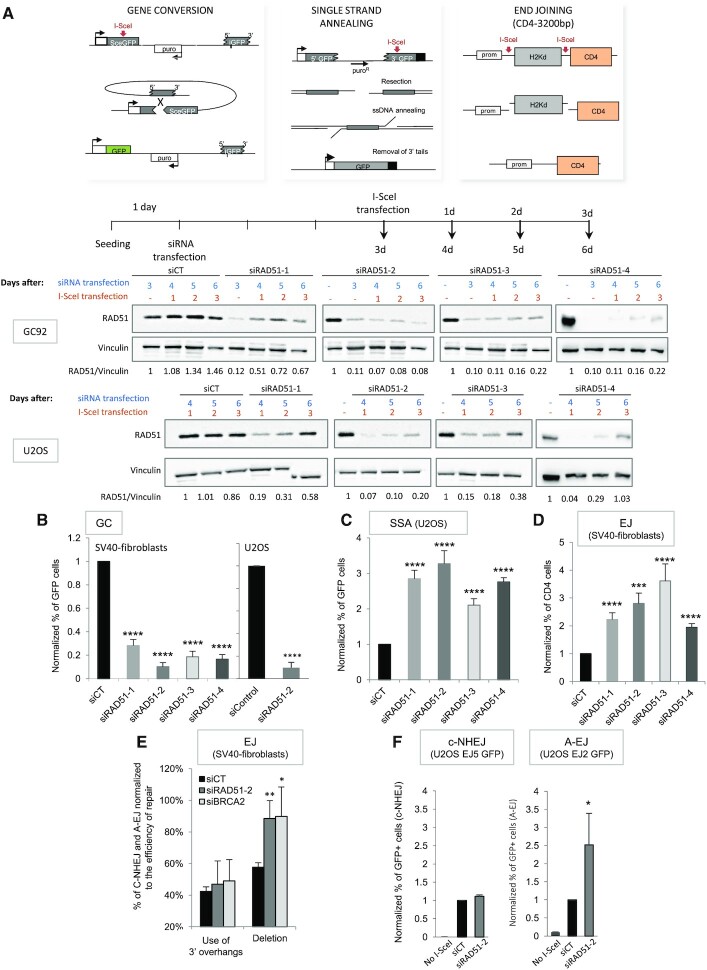
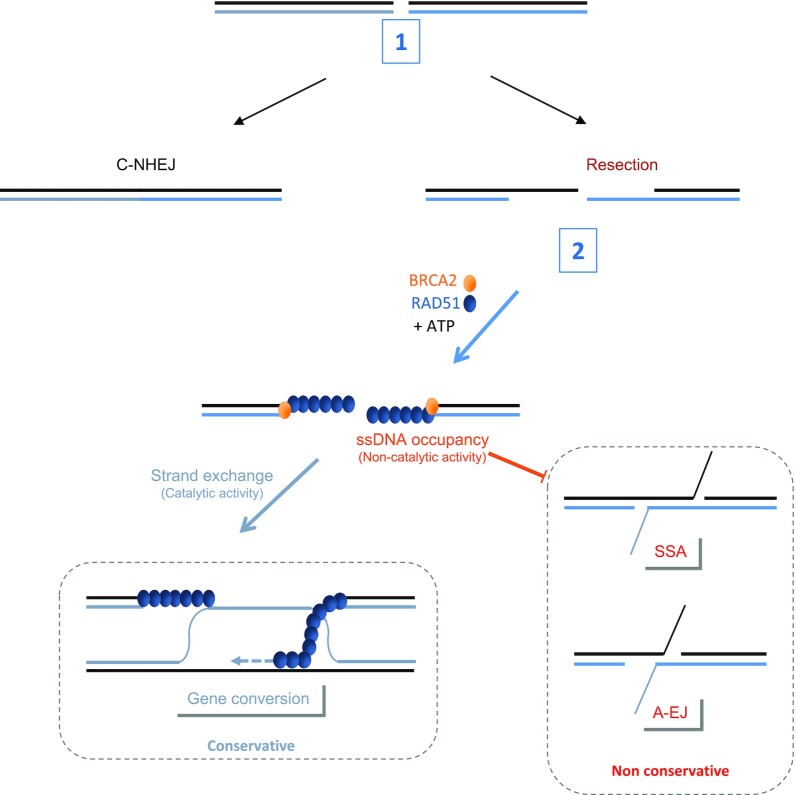
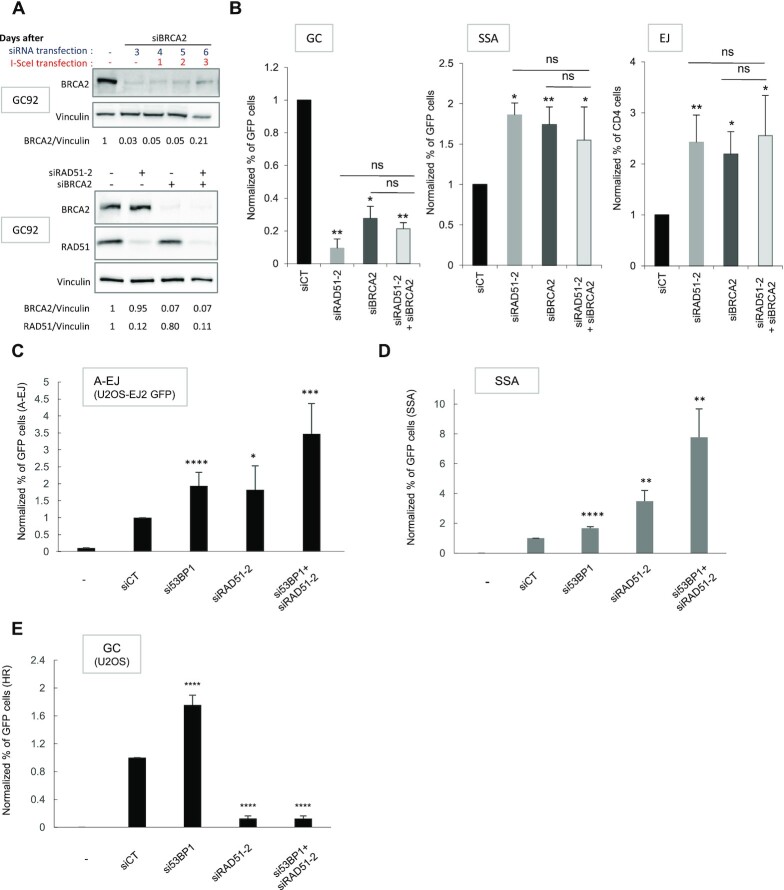
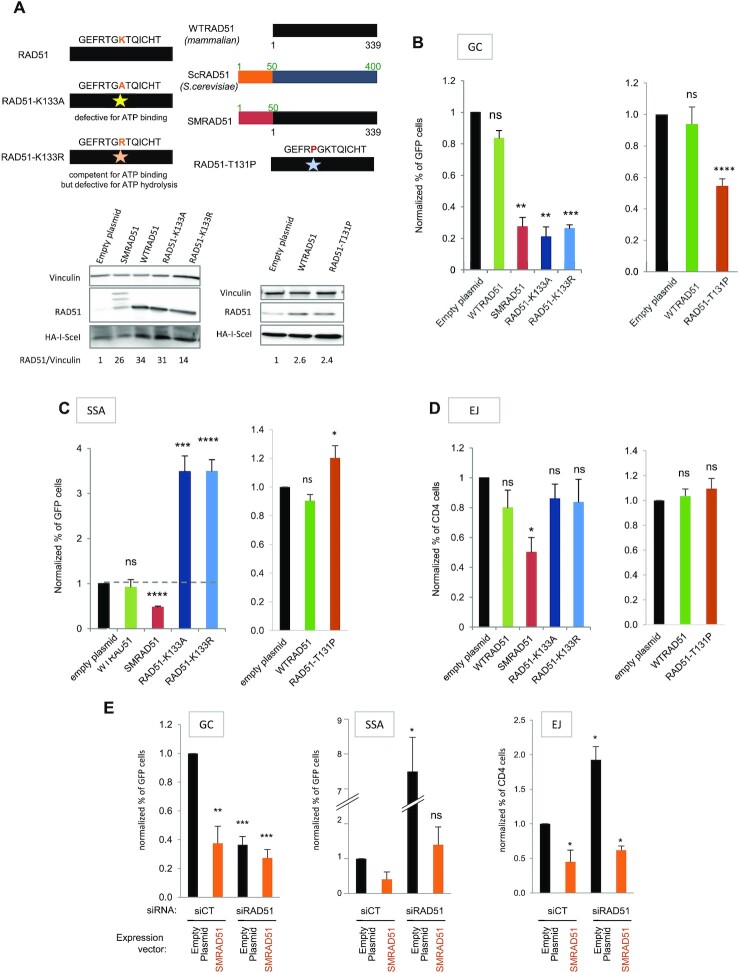
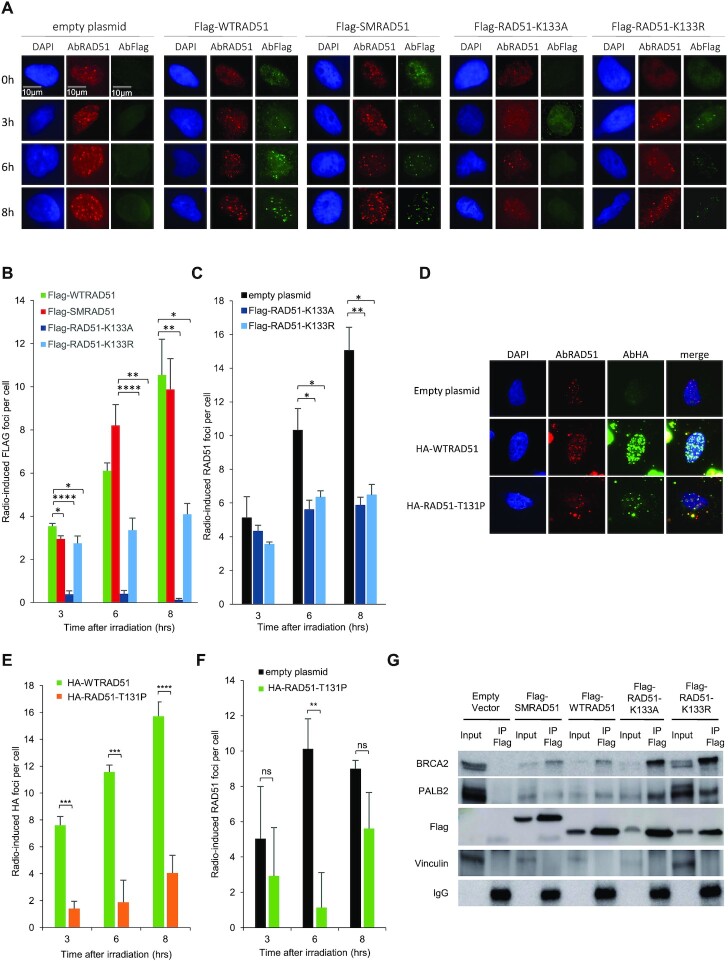
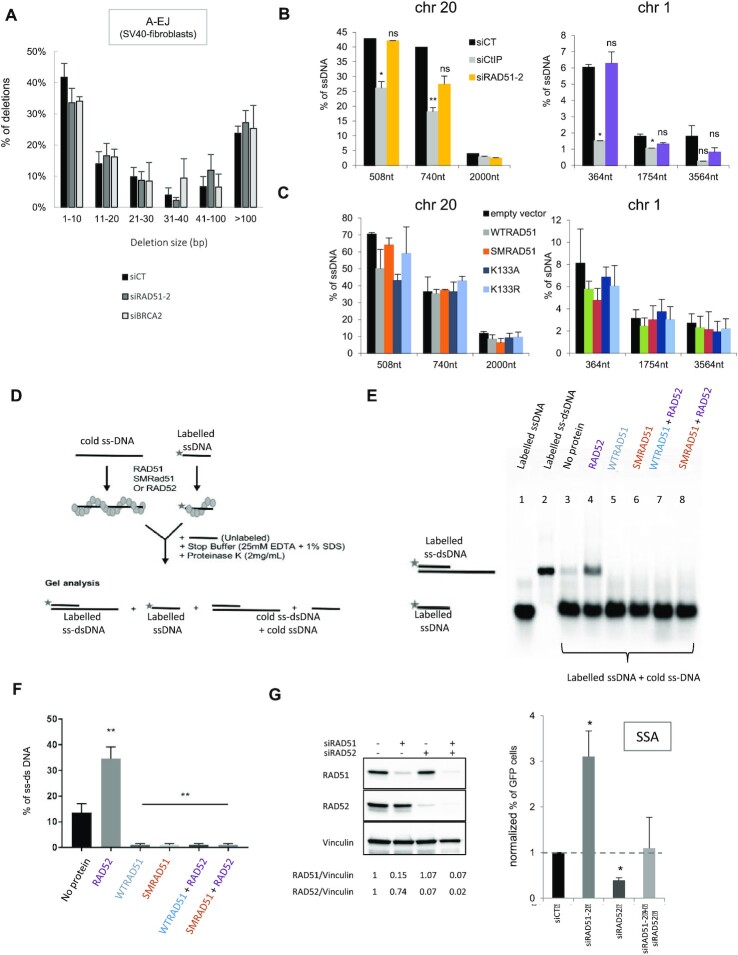
Selection of the appropriate DNA double-strand break (DSB) repair pathway is decisive for genetic stability. It is proposed to act according to two steps: 1-canonical nonhomologous end-joining (C-NHEJ) versus resection that generates single-stranded DNA (ssDNA) stretches; 2-on ssDNA, gene conversion (GC) versus nonconservative single-strand annealing (SSA) or alternative end-joining (A-EJ). Here, we addressed the mechanisms by which RAD51 regulates this second step, preventing nonconservative repair in human cells. Silencing RAD51 or BRCA2 stimulated both SSA and A-EJ, but not C-NHEJ, validating the two-step model. Three different RAD51 dominant-negative forms (DN-RAD51s) repressed GC and stimulated SSA/A-EJ. However, a fourth DN-RAD51 repressed SSA/A-EJ, although it efficiently represses GC. In living cells, the three DN-RAD51s that stimulate SSA/A-EJ failed to load efficiently onto damaged chromatin and inhibited the binding of endogenous RAD51, while the fourth DN-RAD51, which inhibits SSA/A-EJ, efficiently loads on damaged chromatin. Therefore, the binding of RAD51 to DNA, rather than its ability to promote GC, is required for SSA/A-EJ inhibition by RAD51. We showed that RAD51 did not limit resection of endonuclease-induced DSBs, but prevented spontaneous and RAD52-induced annealing of complementary ssDNA in vitro. Therefore, RAD51 controls the selection of the DSB repair pathway, protecting genome integrity from nonconservative DSB repair through ssDNA occupancy, independently of the promotion of CG.
选择合适的 DNA 双链断裂 (DSB) 修复途径对遗传稳定性至关重要。它被提议分为两步进行:1-经典非同源末端连接 (C-NHEJ) 与产生单链 DNA (ssDNA) 链的切除;2-在 ssDNA 上,基因转换 (GC) 与非保守单链退火 (SSA) 或替代末端连接 (A-EJ)。在这里,我们研究了 RAD51 调节这第二步的机制,防止人类细胞中非保守修复。沉默 RAD51 或 BRCA2 刺激 SSA 和 A-EJ,但不刺激 C-NHEJ,验证了两步模型。三种不同的 RAD51 显性失活形式 (DN-RAD51) 抑制 GC 并刺激 SSA/A-EJ。然而,第四种 DN-RAD51 抑制 SSA/A-EJ,尽管它有效地抑制 GC。在活细胞中,三种刺激 SSA/A-EJ 的 DN-RAD51 不能有效地加载到受损染色质上,并抑制内源性 RAD51 的结合,而抑制 SSA/A-EJ 的第四种 DN-RAD51 则有效地加载到受损染色质上。因此,RAD51 与 DNA 的结合,而不是其促进 GC 的能力,是 RAD51 抑制 SSA/A-EJ 的必要条件。我们表明,RAD51 并不限制内切酶诱导的 DSB 切除,但可防止自发和 RAD52 诱导的互补 ssDNA 在体外退火。因此,RAD51 通过 ssDNA 占据控制 DSB 修复途径的选择,保护基因组完整性免受非保守 DSB 修复,而不依赖于 CG 的促进。