Center for Genomic Integrity, Institute for Basic Science, Ulsan, Korea.
Interdisciplinary Graduate Program in Biophysics and Chemical Biology, Seoul National University, Seoul, 08826, Republic of Korea.
Nat Commun. 2019 Dec 16;10(1):5718. doi: 10.1038/s41467-019-13667-4.
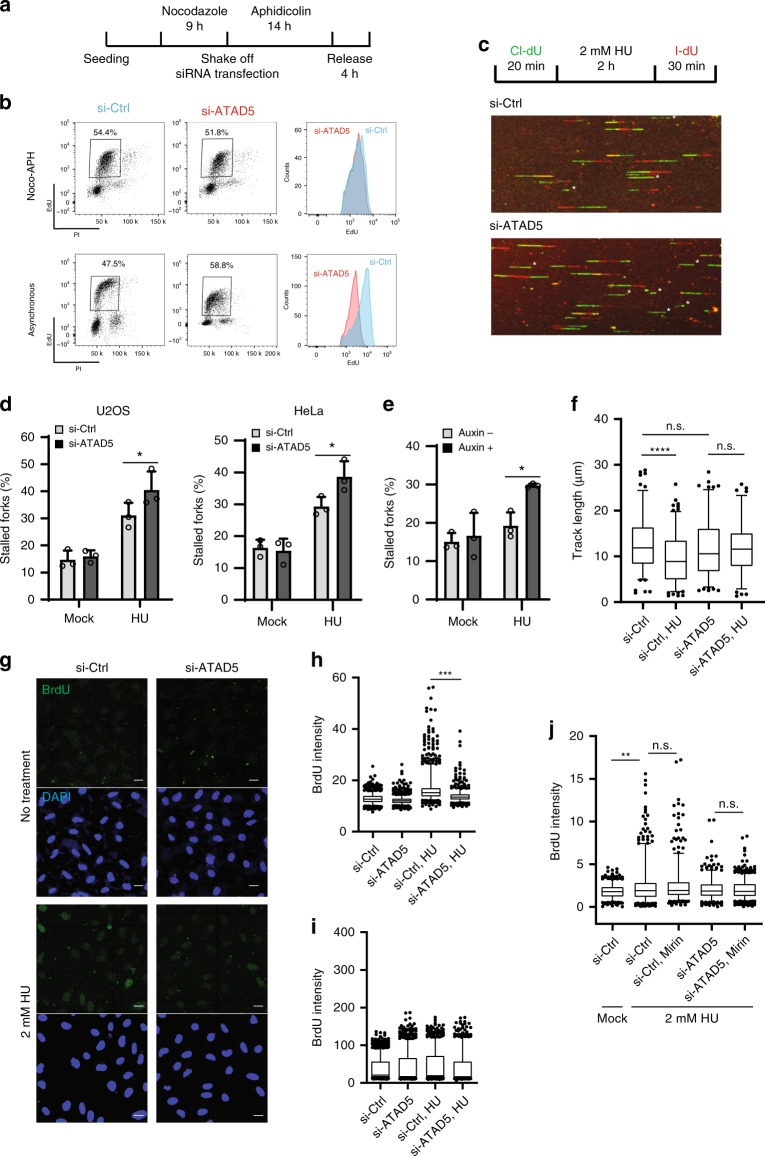
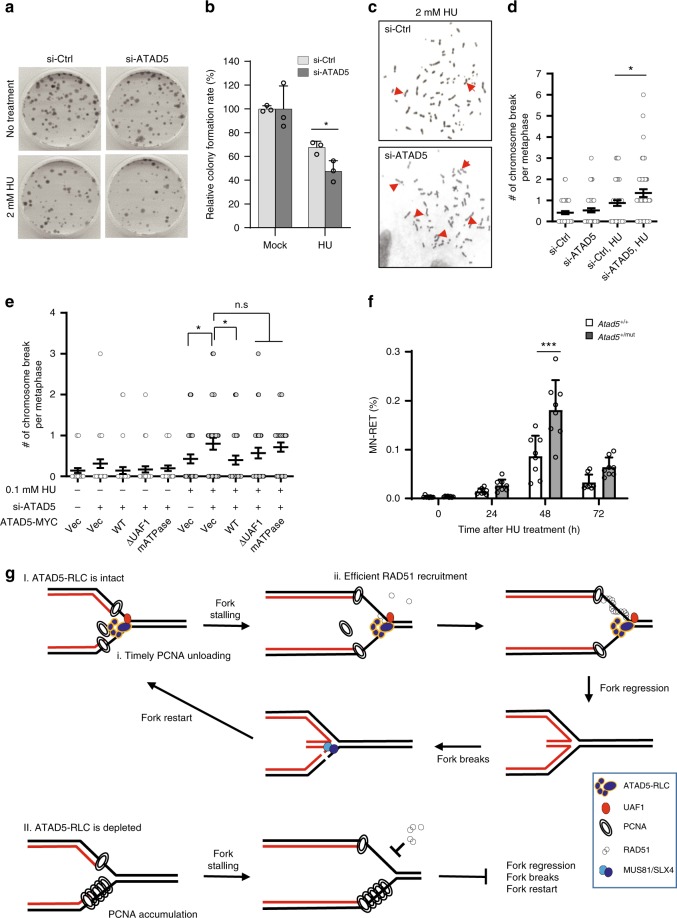
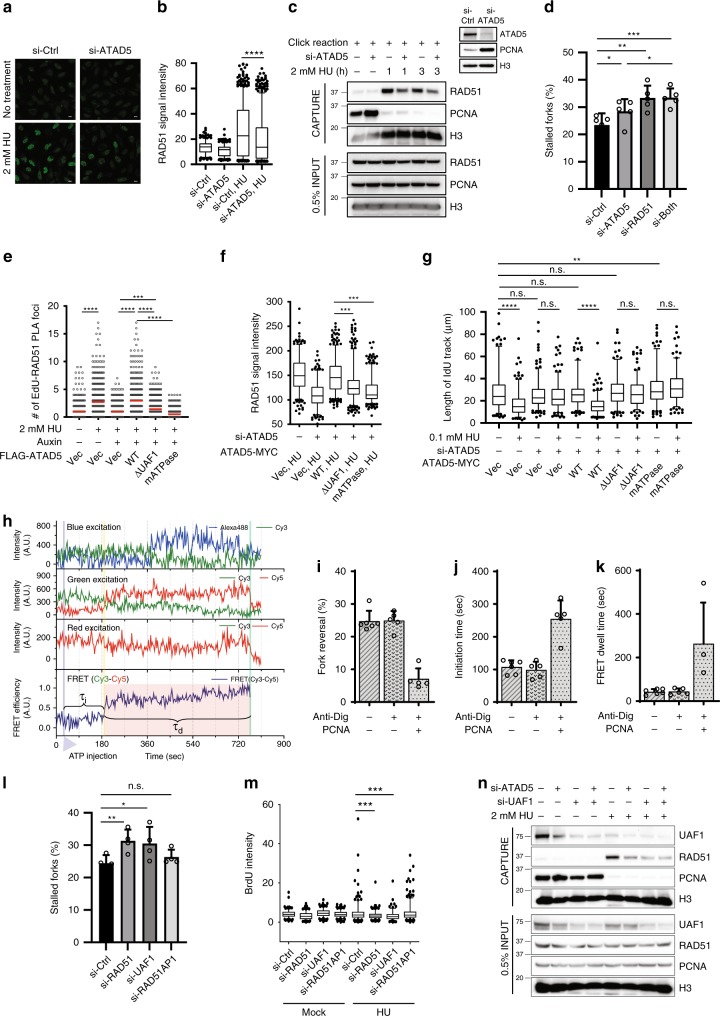
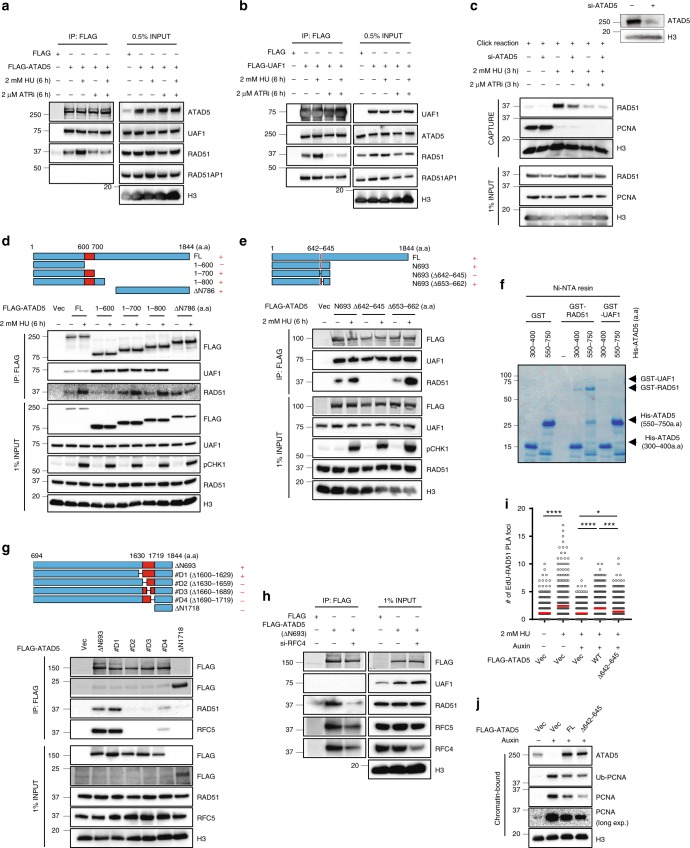
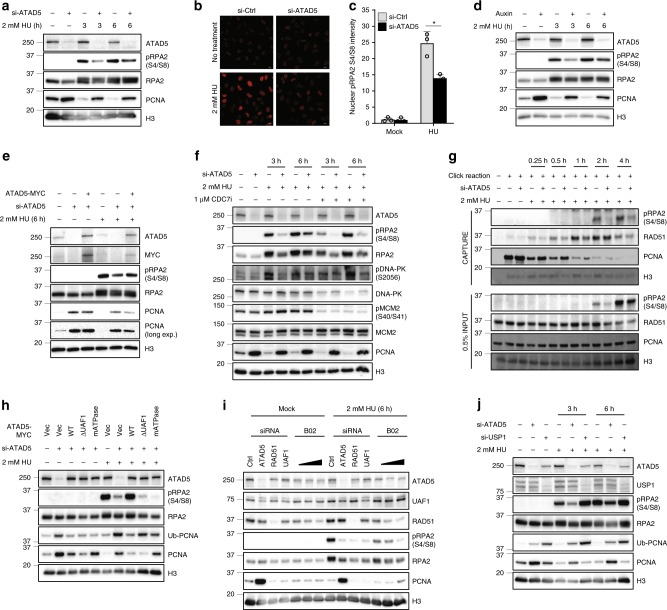
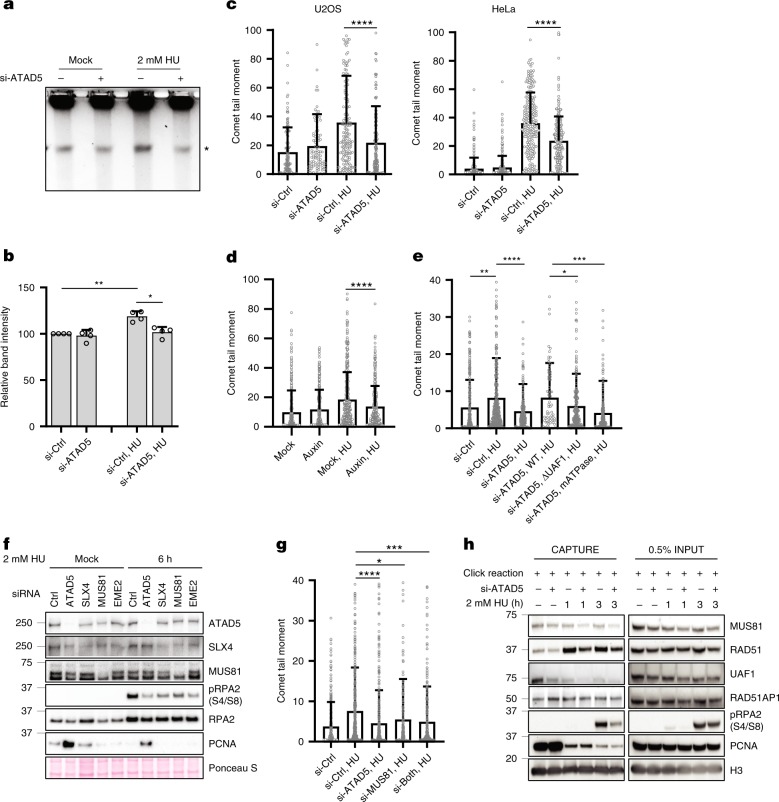
Maintaining stability of replication forks is important for genomic integrity. However, it is not clear how replisome proteins contribute to fork stability under replication stress. Here, we report that ATAD5, a PCNA unloader, plays multiple functions at stalled forks including promoting its restart. ATAD5 depletion increases genomic instability upon hydroxyurea treatment in cultured cells and mice. ATAD5 recruits RAD51 to stalled forks in an ATR kinase-dependent manner by hydroxyurea-enhanced protein-protein interactions and timely removes PCNA from stalled forks for RAD51 recruitment. Consistent with the role of RAD51 in fork regression, ATAD5 depletion inhibits slowdown of fork progression and native 5-bromo-2'-deoxyuridine signal induced by hydroxyurea. Single-molecule FRET showed that PCNA itself acts as a mechanical barrier to fork regression. Consequently, DNA breaks required for fork restart are reduced by ATAD5 depletion. Collectively, our results suggest an important role of ATAD5 in maintaining genome integrity during replication stress.
维持复制叉的稳定性对于基因组完整性很重要。然而,目前尚不清楚复制体蛋白在复制应激下如何有助于维持叉的稳定性。在这里,我们报告说,PCNA 卸载蛋白 ATAD5 在停滞的叉上发挥多种功能,包括促进其重新启动。在培养细胞和小鼠中,羟基脲处理后,ATAD5 耗竭会增加基因组不稳定性。ATAD5 通过羟基脲增强的蛋白-蛋白相互作用,以 ATR 激酶依赖性的方式将 RAD51 募集到停滞的叉上,并及时将 PCNA 从停滞的叉上移除,以招募 RAD51。与 RAD51 在叉回归中的作用一致,ATAD5 耗竭抑制了羟基脲诱导的叉前进速度减慢和天然 5-溴-2'-脱氧尿苷信号。单分子 FRET 表明,PCNA 本身充当叉回归的机械屏障。因此,通过 ATAD5 耗竭减少了叉重新启动所需的 DNA 断裂。总之,我们的结果表明 ATAD5 在复制应激期间维持基因组完整性方面起着重要作用。