Department of Biological Sciences, Vanderbilt University, Nashville, Tennessee, United States of America.
PLoS Genet. 2020 Feb 3;16(2):e1008608. doi: 10.1371/journal.pgen.1008608. eCollection 2020 Feb.
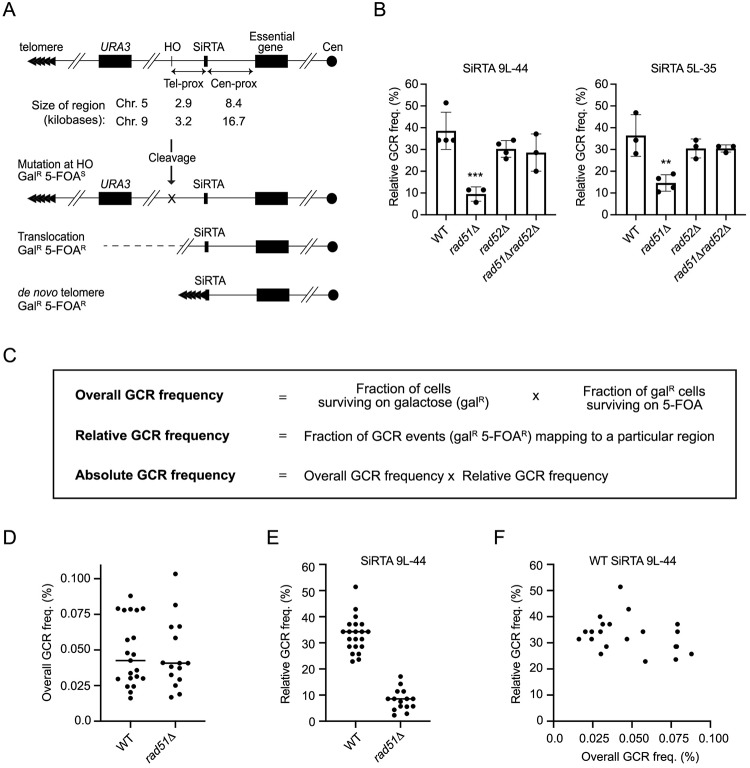
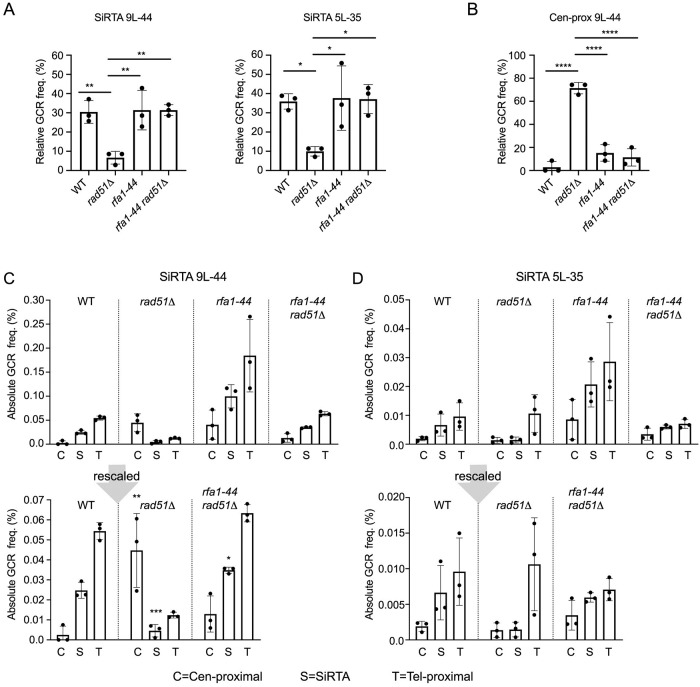
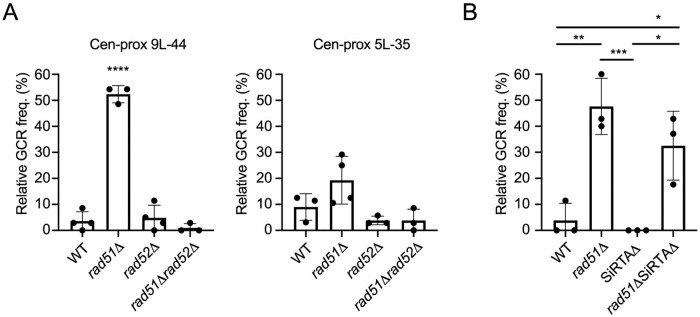
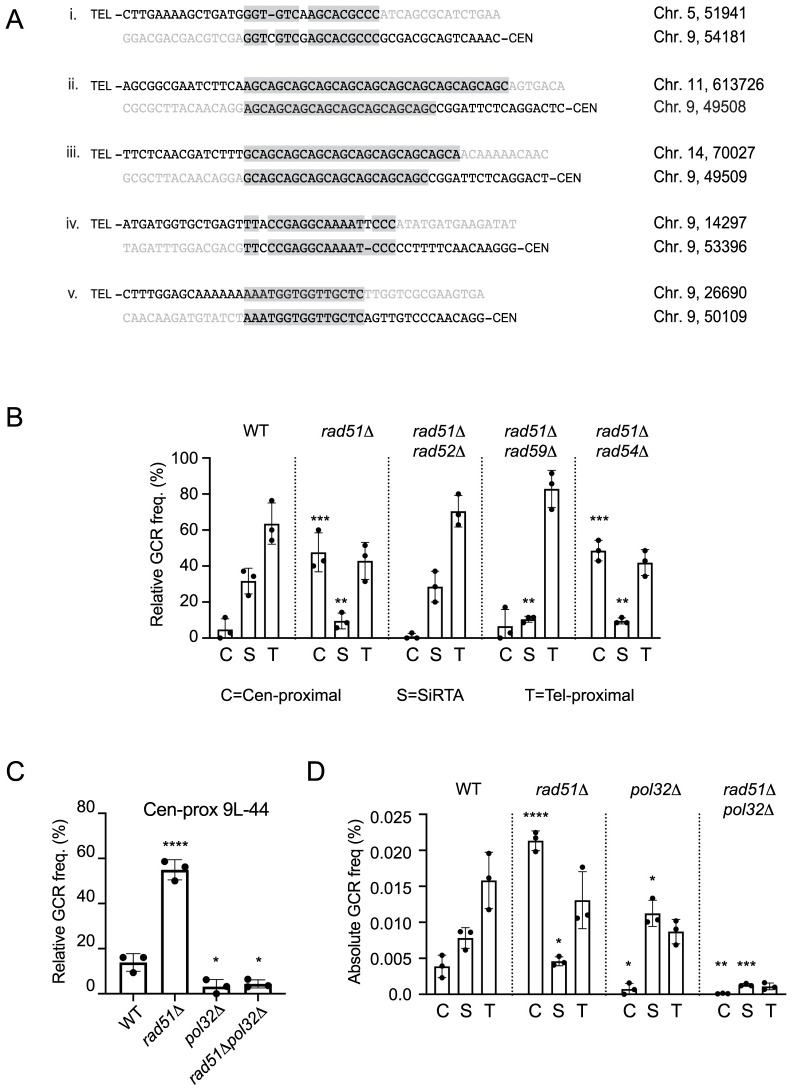
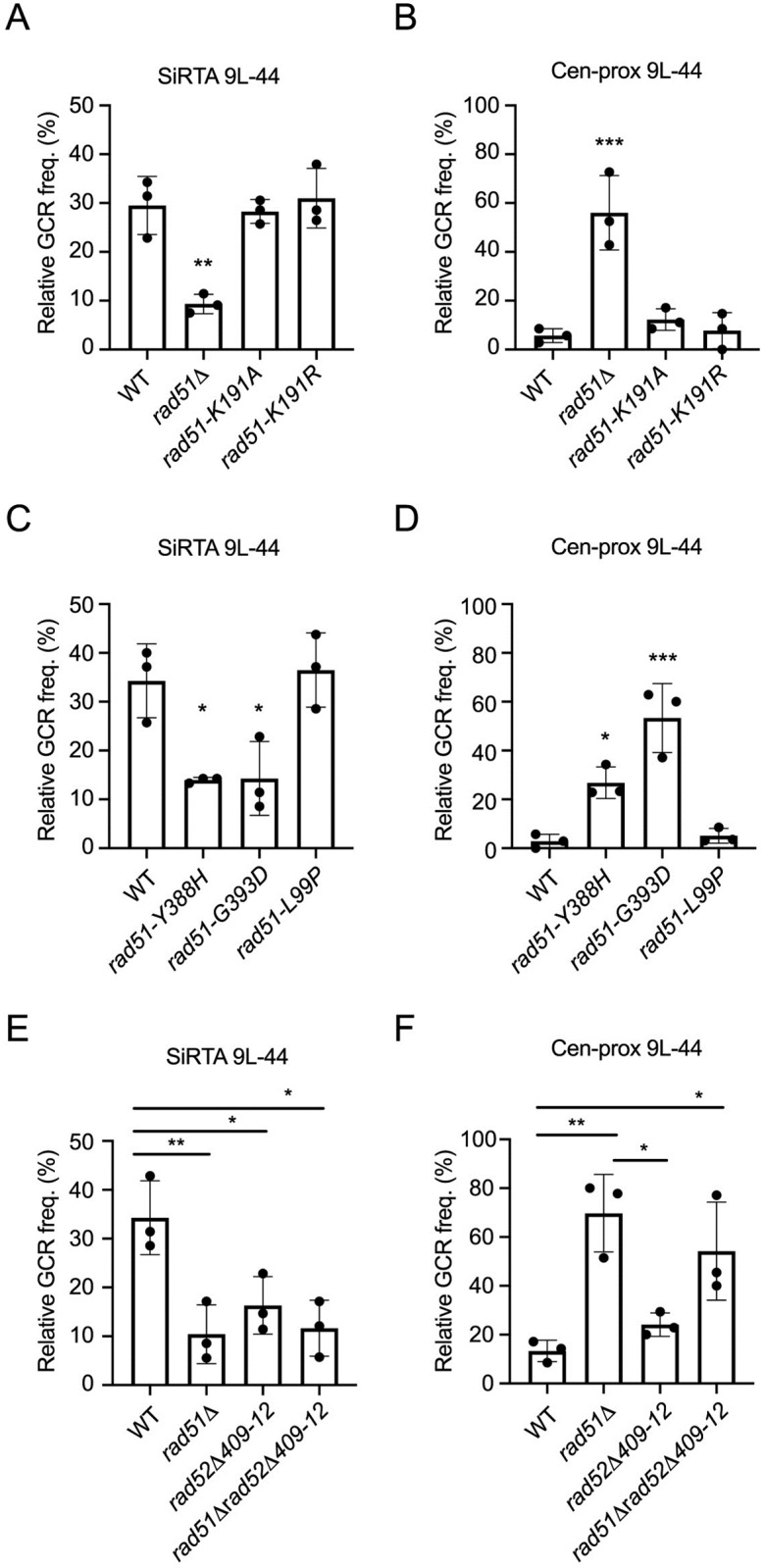
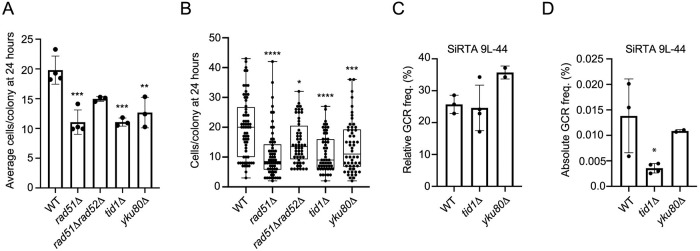
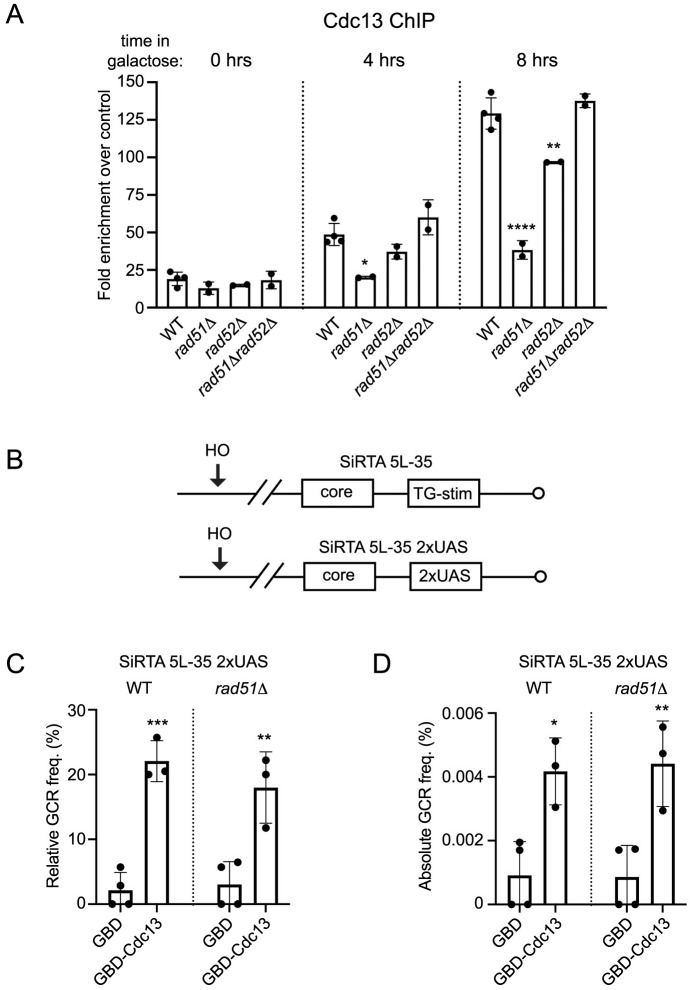
DNA double-strand breaks (DSBs) are toxic forms of DNA damage that must be repaired to maintain genome integrity. Telomerase can act upon a DSB to create a de novo telomere, a process that interferes with normal repair and creates terminal deletions. We previously identified sequences in Saccharomyces cerevisiae (SiRTAs; Sites of Repair-associated Telomere Addition) that undergo unusually high frequencies of de novo telomere addition, even when the original chromosome break is several kilobases distal to the eventual site of telomerase action. Association of the single-stranded telomere binding protein Cdc13 with a SiRTA is required to stimulate de novo telomere addition. Because extensive resection must occur prior to Cdc13 binding, we utilized these sites to monitor the effect of proteins involved in homologous recombination. We find that telomere addition is significantly reduced in the absence of the Rad51 recombinase, while loss of Rad52, required for Rad51 nucleoprotein filament formation, has no effect. Deletion of RAD52 suppresses the defect of the rad51Δ strain, suggesting that Rad52 inhibits de novo telomere addition in the absence of Rad51. The ability of Rad51 to counteract this effect of Rad52 does not require DNA binding by Rad51, but does require interaction between the two proteins, while the inhibitory effect of Rad52 depends on its interaction with Replication Protein A (RPA). Intriguingly, the genetic interactions we report between RAD51 and RAD52 are similar to those previously observed in the context of checkpoint adaptation. Forced recruitment of Cdc13 fully restores telomere addition in the absence of Rad51, suggesting that Rad52, through its interaction with RPA-coated single-stranded DNA, inhibits the ability of Cdc13 to bind and stimulate telomere addition. Loss of the Rad51-Rad52 interaction also stimulates a subset of Rad52-dependent microhomology-mediated repair (MHMR) events, consistent with the known ability of Rad51 to prevent single-strand annealing.
DNA 双链断裂 (DSBs) 是一种有毒的 DNA 损伤形式,必须进行修复以维持基因组完整性。端粒酶可以作用于 DSB 来创建新的端粒,这一过程干扰了正常的修复并产生末端缺失。我们之前在酿酒酵母中鉴定了一些序列(SiRTAs;修复相关端粒添加位点),即使原始染色体断裂距离端粒酶作用的最终位点有几千个碱基对,它们也会经历异常高的新端粒添加频率。单链端粒结合蛋白 Cdc13 与 SiRTA 的结合对于刺激新端粒的添加是必需的。由于在 Cdc13 结合之前必须发生广泛的切除,因此我们利用这些位点来监测参与同源重组的蛋白质的影响。我们发现,在缺乏 Rad51 重组酶的情况下,端粒添加显著减少,而 Rad52 的缺失(Rad51 核蛋白丝形成所必需的)则没有影响。RAD52 的缺失抑制了 rad51Δ 菌株的缺陷,表明 Rad52 在没有 Rad51 的情况下抑制新端粒的添加。Rad51 抵消 Rad52 这种效应的能力不需要 Rad51 的 DNA 结合,但确实需要两种蛋白质之间的相互作用,而 Rad52 的抑制作用依赖于其与复制蛋白 A (RPA) 的相互作用。有趣的是,我们报告的 RAD51 和 RAD52 之间的遗传相互作用与之前在检查点适应背景下观察到的相互作用相似。Cdc13 的强制募集在缺乏 Rad51 的情况下完全恢复了端粒的添加,这表明 Rad52 通过与 RPA 包裹的单链 DNA 的相互作用,抑制了 Cdc13 结合和刺激端粒添加的能力。Rad51-Rad52 相互作用的丧失也刺激了一组 Rad52 依赖性微同源介导修复 (MHMR) 事件,这与已知的 Rad51 防止单链退火的能力一致。