Institute of Molecular Health Sciences, ETH Zürich, Zürich, Switzerland.
Elife. 2020 Feb 25;9:e53627. doi: 10.7554/eLife.53627.
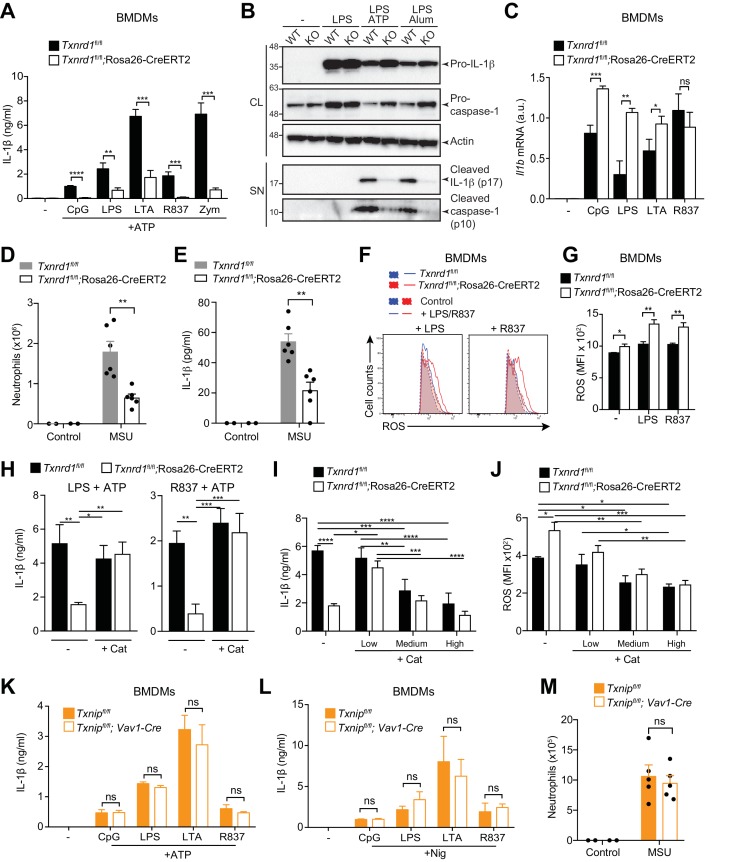
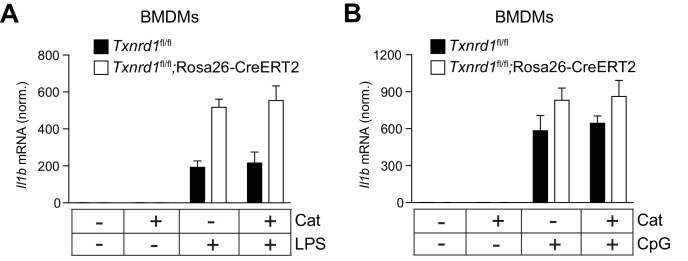
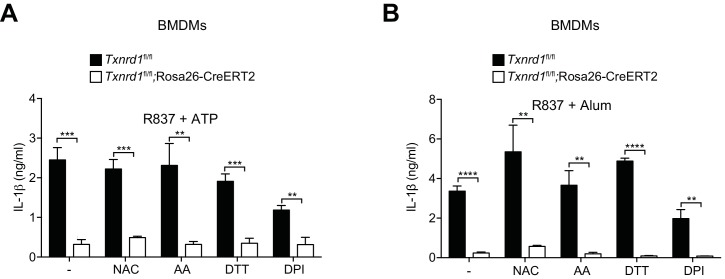
Antioxidant systems, such as the thioredoxin-1 (Trx1) pathway, ensure cellular redox homeostasis. However, how such systems regulate development and function of myeloid cells is barely understood. Here we show that in contrast to its critical role in T cells, the murine Trx1 system is dispensable for steady-state myeloid-cell hematopoiesis due to their capacity to tap the glutathione/glutaredoxin pathway for DNA biosynthesis. However, the Trx1 pathway instrumentally enables nuclear NF-κB DNA-binding and thereby pro-inflammatory responses in monocytes and dendritic cells. Moreover, independent of this activity, Trx1 is critical for NLRP3 inflammasome activation and IL-1β production in macrophages by detoxifying excessive ROS levels. Notably, we exclude the involvement of the Trx1 inhibitor Txnip as a redox-sensitive ligand of NLRP3 as previously proposed. Together, this study suggests that targeting Trx1 may be exploited to treat inflammatory diseases.
抗氧化系统,如硫氧还蛋白-1 (Trx1) 途径,可确保细胞的氧化还原平衡。然而,人们对这些系统如何调节髓样细胞的发育和功能知之甚少。在这里,我们表明,与在 T 细胞中的关键作用相反,由于其能够利用谷胱甘肽/谷胱甘肽还原酶途径进行 DNA 合成,因此小鼠的 Trx1 系统对于稳态髓样细胞造血是可有可无的。然而,Trx1 途径有助于单核细胞和树突状细胞中的核 NF-κB DNA 结合,从而引发炎症反应。此外,Trx1 对于巨噬细胞中的 NLRP3 炎性小体激活和 IL-1β 产生是至关重要的,它可清除过多的 ROS 水平。值得注意的是,我们排除了 Trx1 抑制剂 Txnip 作为 NLRP3 的氧化还原敏感配体的参与,正如之前所提出的那样。总之,这项研究表明,靶向 Trx1 可能被用于治疗炎症性疾病。