Department of Chemistry and Biochemistry, University of California Santa Cruz, Santa Cruz, United States.
Institute of Transformative Bio-Molecules, Nagoya University, Nagoya, Japan.
Elife. 2020 Feb 26;9:e55275. doi: 10.7554/eLife.55275.
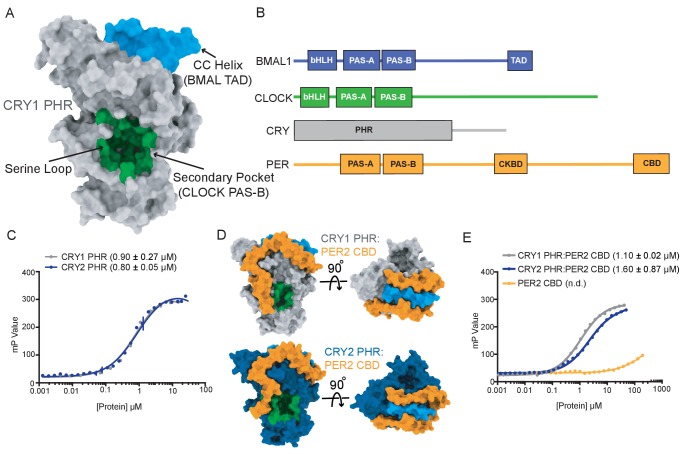
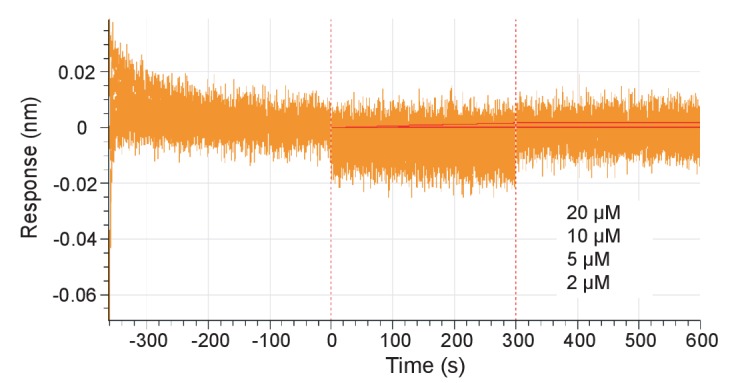
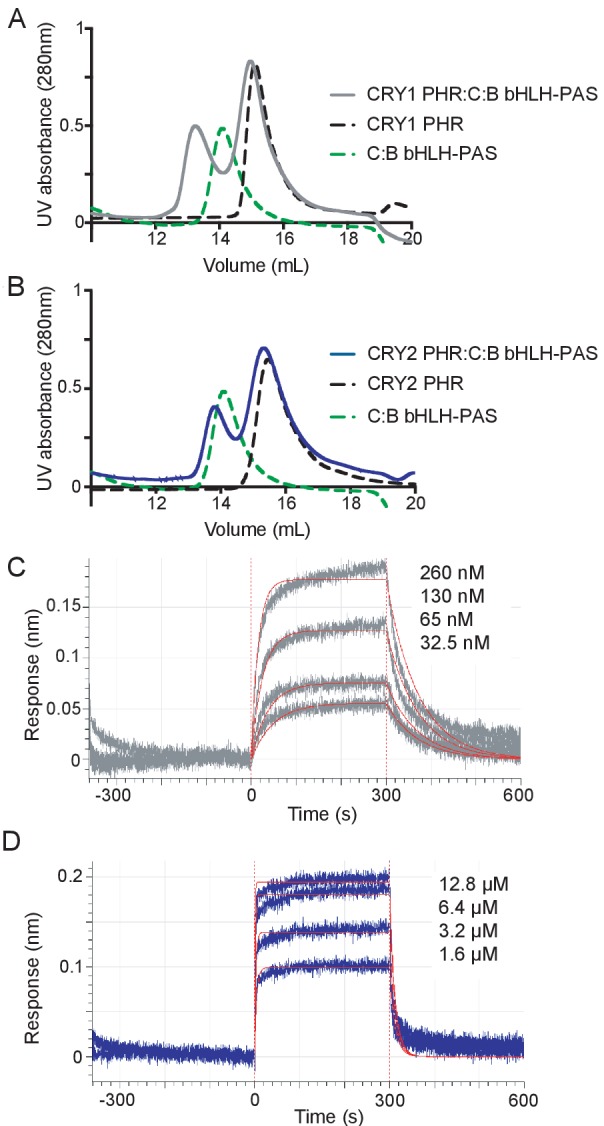
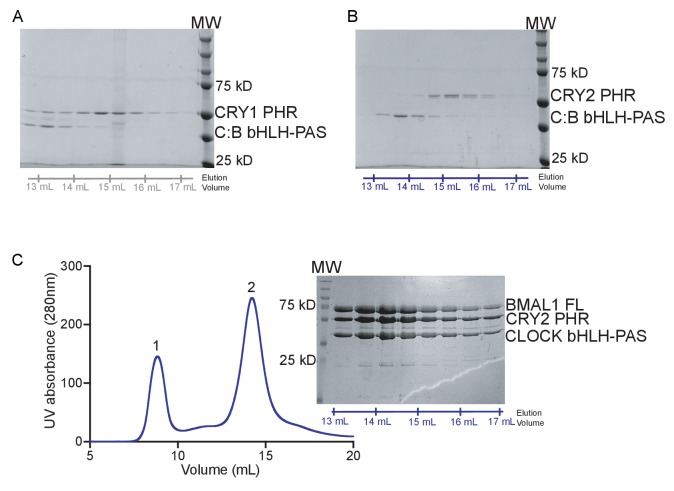
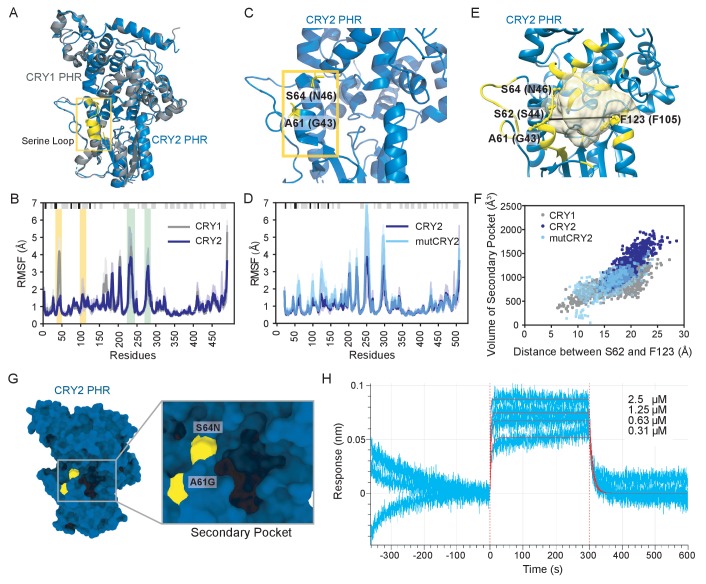
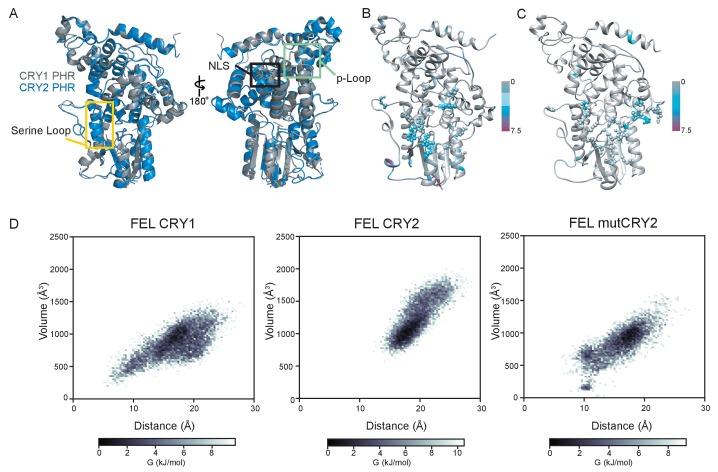
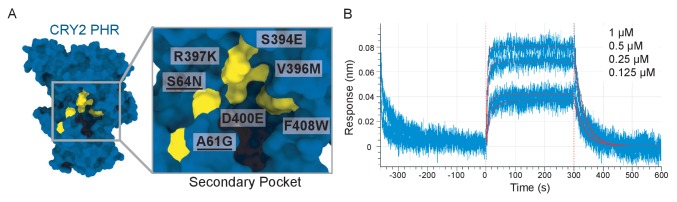
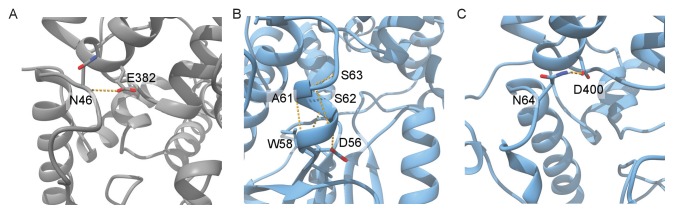
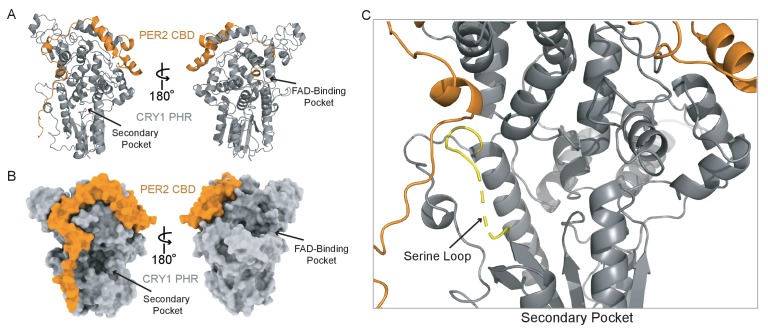
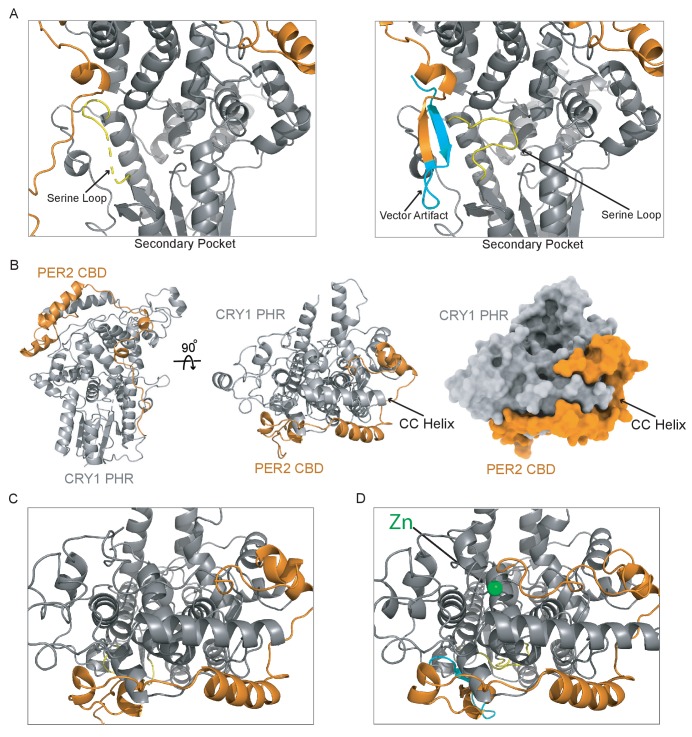
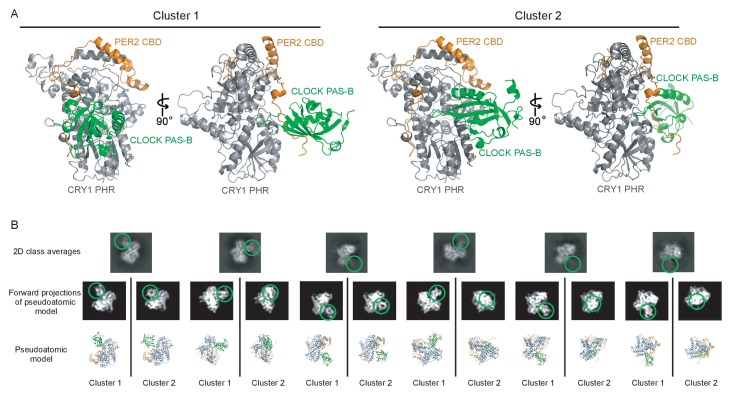
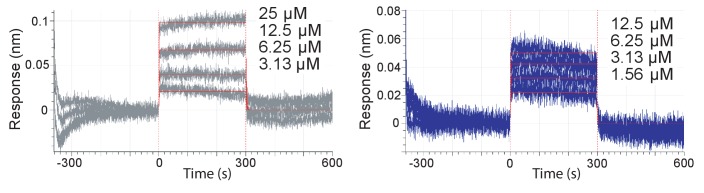
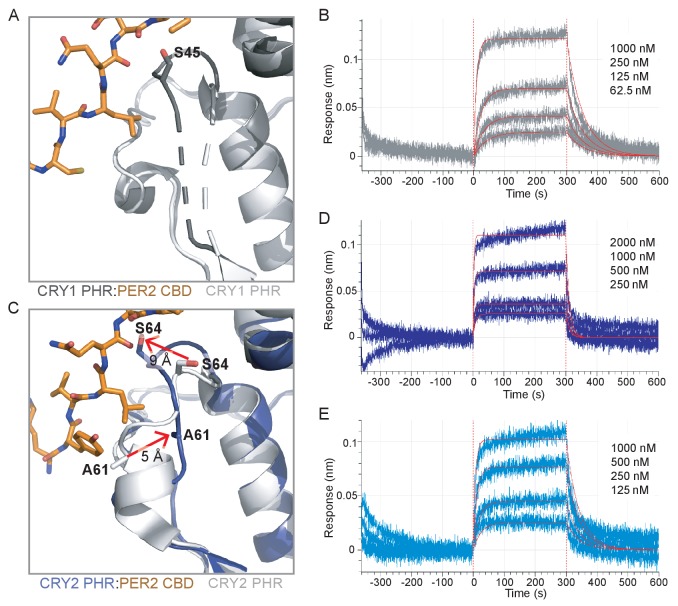
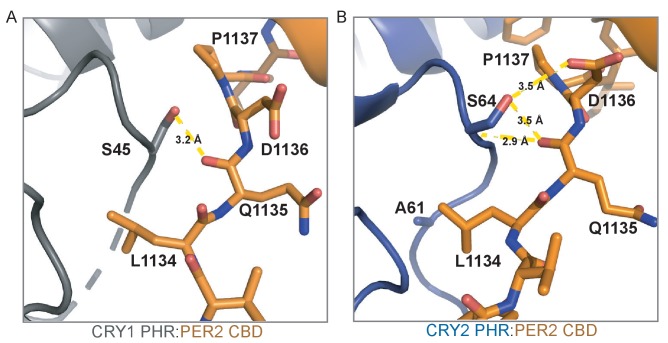
Mammalian circadian rhythms are generated by a transcription-based feedback loop in which CLOCK:BMAL1 drives transcription of its repressors (PER1/2, CRY1/2), which ultimately interact with CLOCK:BMAL1 to close the feedback loop with ~24 hr periodicity. Here we pinpoint a key difference between CRY1 and CRY2 that underlies their differential strengths as transcriptional repressors. Both cryptochromes bind the BMAL1 transactivation domain similarly to sequester it from coactivators and repress CLOCK:BMAL1 activity. However, we find that CRY1 is recruited with much higher affinity to the PAS domain core of CLOCK:BMAL1, allowing it to serve as a stronger repressor that lengthens circadian period. We discovered a dynamic serine-rich loop adjacent to the secondary pocket in the photolyase homology region (PHR) domain that regulates differential binding of cryptochromes to the PAS domain core of CLOCK:BMAL1. Notably, binding of the co-repressor PER2 remodels the serine loop of CRY2, making it more CRY1-like and enhancing its affinity for CLOCK:BMAL1.
哺乳动物的昼夜节律是由一个基于转录的反馈环产生的,其中 CLOCK:BMAL1 驱动其抑制剂(PER1/2、CRY1/2)的转录,这些抑制剂最终与 CLOCK:BMAL1 相互作用,以约 24 小时的周期性关闭反馈环。在这里,我们发现 CRY1 和 CRY2 之间的一个关键区别,这是它们作为转录抑制剂的差异强度的基础。两个隐花色素都以相似的方式结合 BMAL1 的转录激活结构域,将其与共激活因子隔离,并抑制 CLOCK:BMAL1 的活性。然而,我们发现 CRY1 以更高的亲和力被招募到 CLOCK:BMAL1 的 PAS 结构域核心,使其能够作为更强的抑制剂,延长生物钟周期。我们发现一个动态的富含丝氨酸的环,位于光解酶同源区(PHR)域的次要口袋附近,调节隐花色素与 CLOCK:BMAL1 的 PAS 结构域核心的差异结合。值得注意的是,共抑制剂 PER2 结合重塑了 CRY2 的丝氨酸环,使其更类似于 CRY1,并增强了其与 CLOCK:BMAL1 的亲和力。