Department of Molecular Biosciences, Northwestern University, Evanston, United States.
NSF-Simons Center for Quantitative Biology, Northwestern University, Evanston, United States.
Elife. 2020 Feb 26;9:e53638. doi: 10.7554/eLife.53638.
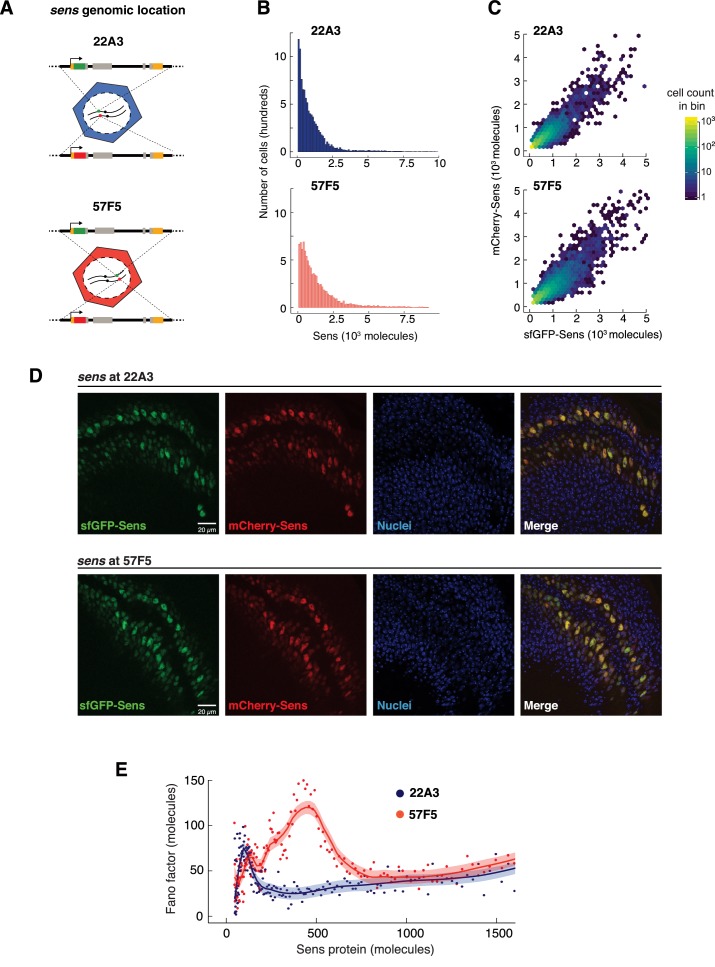
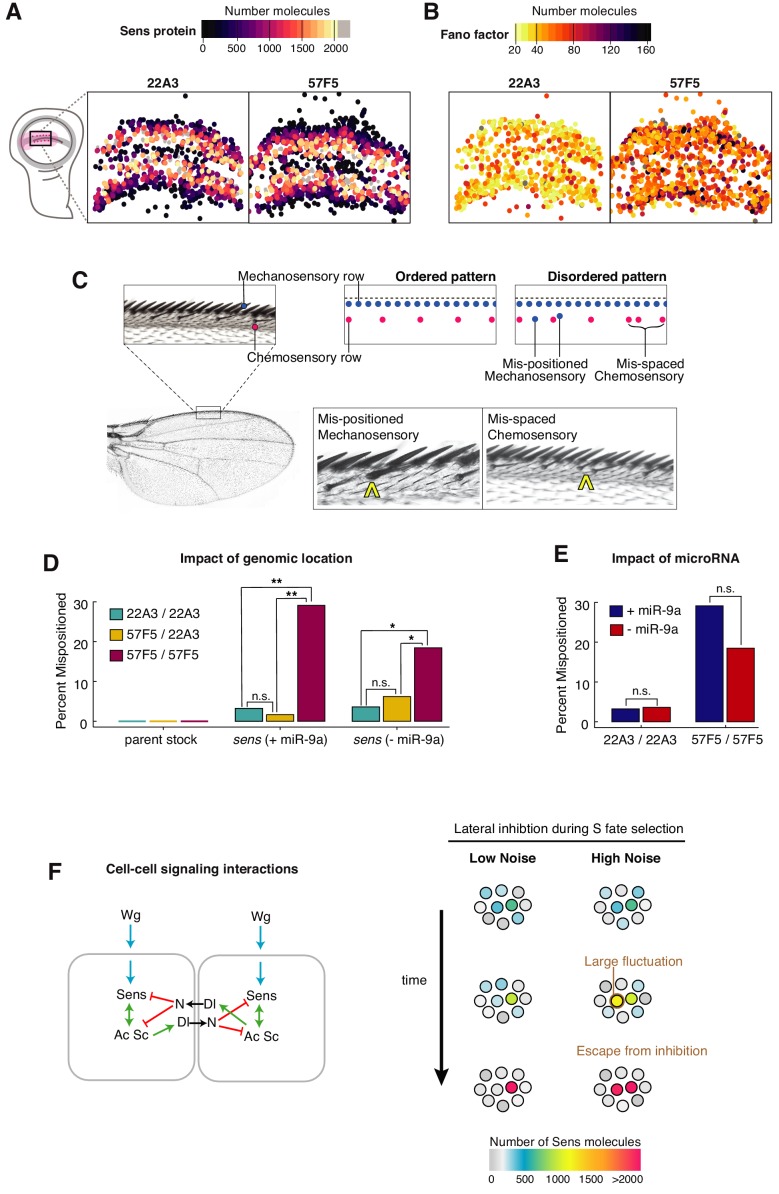
Sensory neuron numbers and positions are precisely organized to accurately map environmental signals in the brain. This precision emerges from biochemical processes within and between cells that are inherently stochastic. We investigated impact of stochastic gene expression on pattern formation, focusing on (), a key determinant of sensory fate in . Perturbing microRNA regulation or genomic location of produced distinct noise signatures. Noise was greatly enhanced when both alleles were present in homologous loci such that each allele was regulated in trans by the other allele. This led to disordered patterning. In contrast, loss of microRNA repression of increased protein abundance but not sensory pattern disorder. This suggests that gene expression stochasticity is a critical feature that must be constrained during development to allow rapid yet accurate cell fate resolution.
感觉神经元的数量和位置经过精确的组织,以在大脑中准确地映射环境信号。这种精确性源自细胞内和细胞间的生化过程,这些过程本质上是随机的。我们研究了随机基因表达对模式形成的影响,重点研究了 (),这是在 () 中决定感觉命运的关键决定因素。干扰 microRNA 调节或 () 的基因组位置会产生不同的噪声特征。当同源基因座中存在两个 () 等位基因时,噪声会大大增强,使得每个等位基因都被另一个等位基因反式调节。这导致了模式的紊乱。相比之下,缺失 microRNA 对 () 的抑制作用会增加蛋白质丰度,但不会导致感觉模式紊乱。这表明基因表达的随机性是一个关键特征,在发育过程中必须受到限制,以允许快速而准确的细胞命运解析。