Era Manami, Matsuo Yosuke, Saito Yoshinori, Tanaka Takashi
Department of Natural Product Chemistry, Graduate School of Biomedical Sciences, Nagasaki University, 1-14 Bunkyo-machi, Nagasaki 852-8521, Japan.
Molecules. 2020 Feb 26;25(5):1051. doi: 10.3390/molecules25051051.
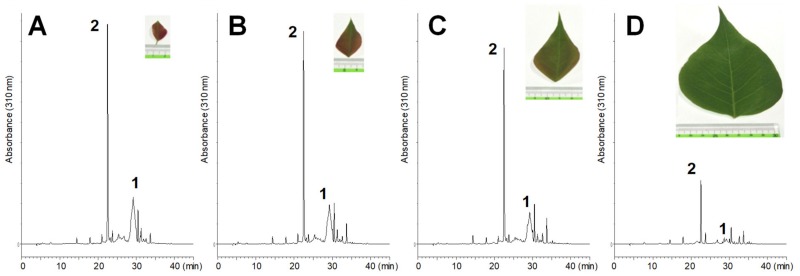
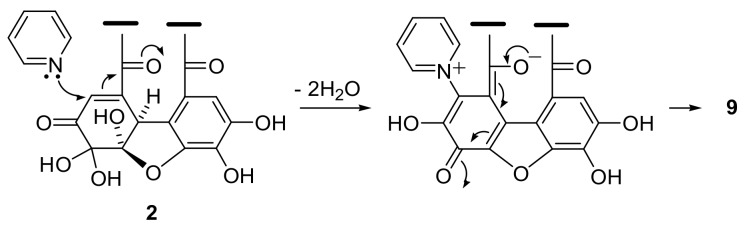
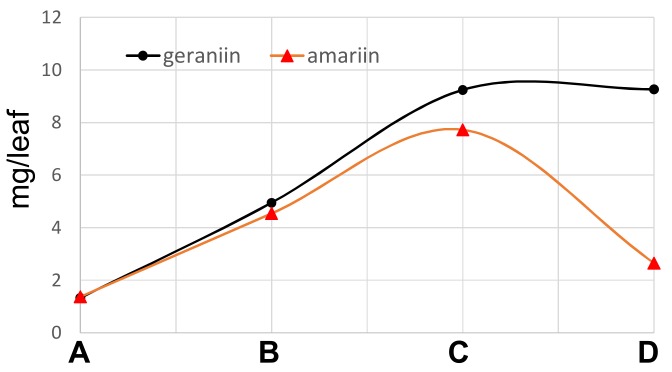
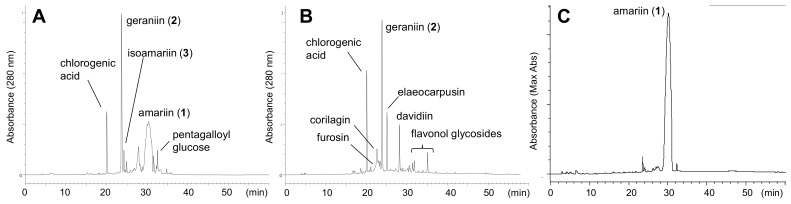
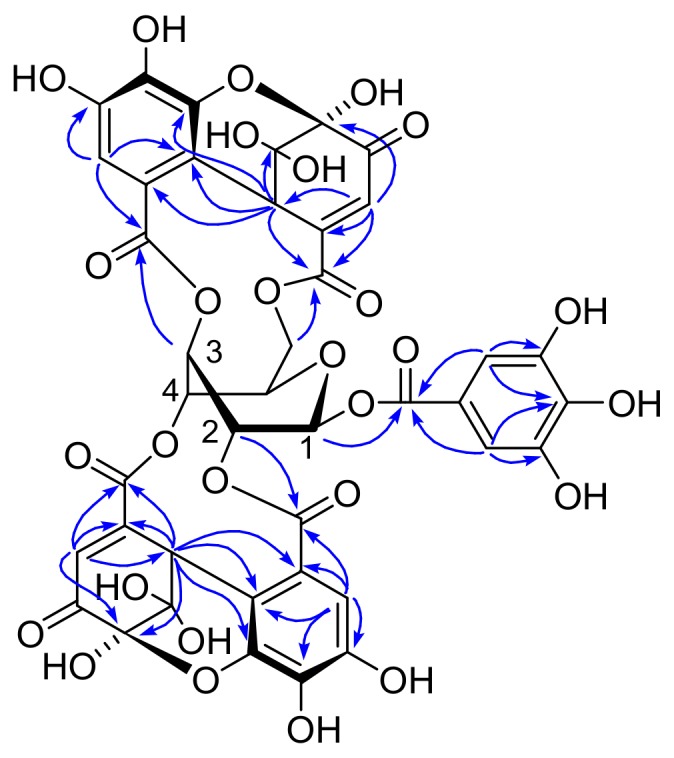
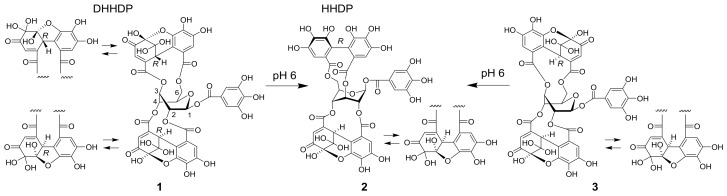
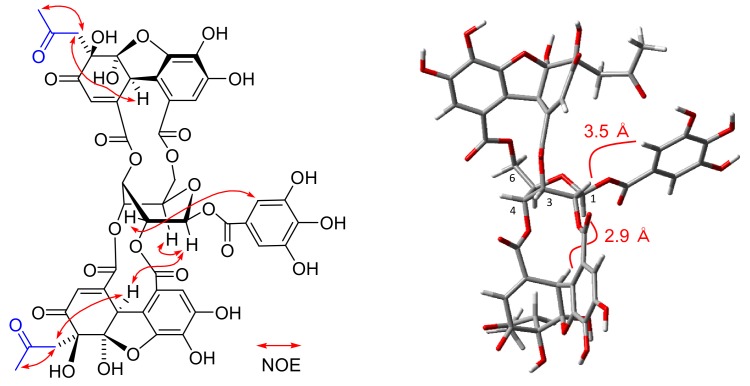
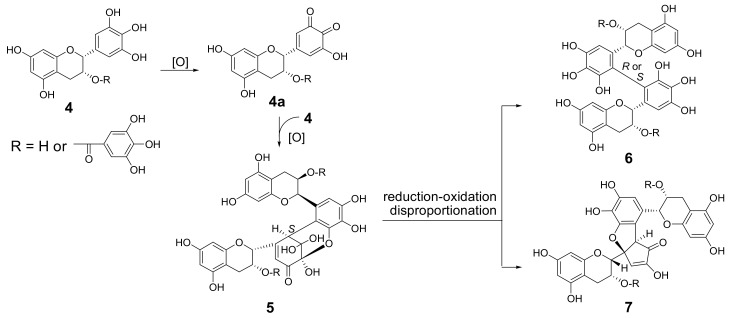
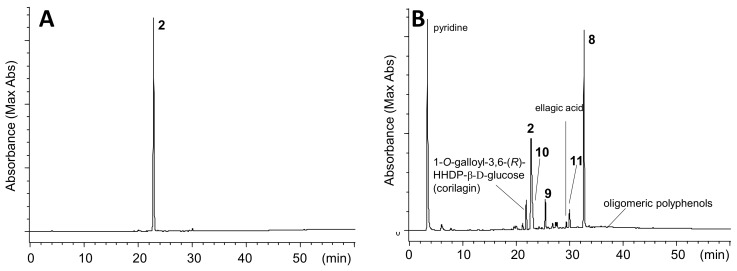
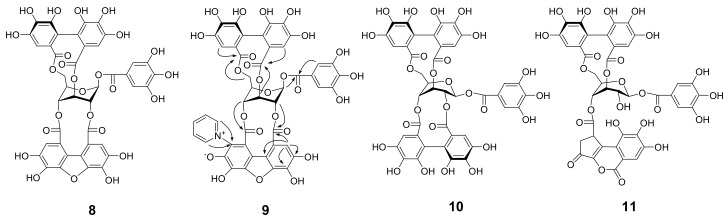
Amariin is an ellagitannin with two dehydrohexahydroxydiphenoyl (DHHDP) moieties connecting glucose 2,4- and 3,6-hydroxy groups. This tannin is predominant in the young leaves of and . However, as the leaves grow, the 3,6-DHHDP is converted to its reduced form, the hexahydroxydiphenoyl (HHDP) group, to generate geraniin, a predominant ellagitannin of the matured leaves. The purified amariin is unstable in aqueous solution, and the 3,6-()-DHHDP is spontaneously degraded to give HHDP, whereas 2,4-()-DHHDP is stable. The driving force of the selective reduction of the 3,6-DHHDP of amariin is shown to be the conformational change of glucose from B to C. Heating geraniin with pyridine affords 2,4-()-DHHDP reduction products. Furthermore, the acid hydrolysis of geraniin yields two equivalents of ellagic acid. Although the reaction mechanism is still ambiguous, these results propose an alternative biosynthetic route of the ellagitannin HHDP groups.
诃子鞣质是一种鞣花单宁,有两个脱氢六羟基二苯甲酰基(DHHDP)部分连接葡萄糖的2,4-和3,6-羟基。这种单宁在[植物名称1]和[植物名称2]的幼叶中占主导地位。然而,随着叶子生长,3,6-DHHDP会转化为其还原形式,即六羟基二苯甲酰基(HHDP)基团,从而生成老鹳草素,这是成熟叶片中占主导地位的鞣花单宁。纯化后的诃子鞣质在水溶液中不稳定,3,6-()-DHHDP会自发降解生成HHDP,而2,4-()-DHHDP则是稳定的。诃子鞣质3,6-DHHDP选择性还原的驱动力被证明是葡萄糖从B型到C型的构象变化。用吡啶加热老鹳草素可得到2,4-()-DHHDP还原产物。此外,老鹳草素的酸水解产生两当量的鞣花酸。尽管反应机制仍不明确,但这些结果提出了鞣花单宁HHDP基团的另一种生物合成途径。