Department of Chemical and Systems Biology, Stanford University School of Medicine, Stanford, United States.
Stanford University Mass Spectrometry, Stanford University, Stanford, United States.
Elife. 2020 Mar 12;9:e50209. doi: 10.7554/eLife.50209.
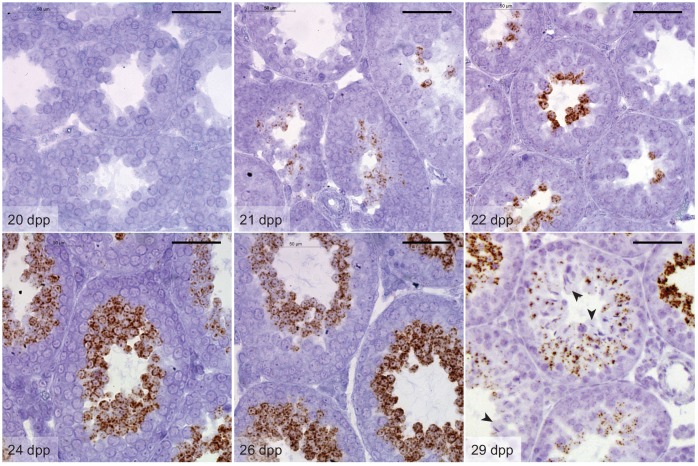
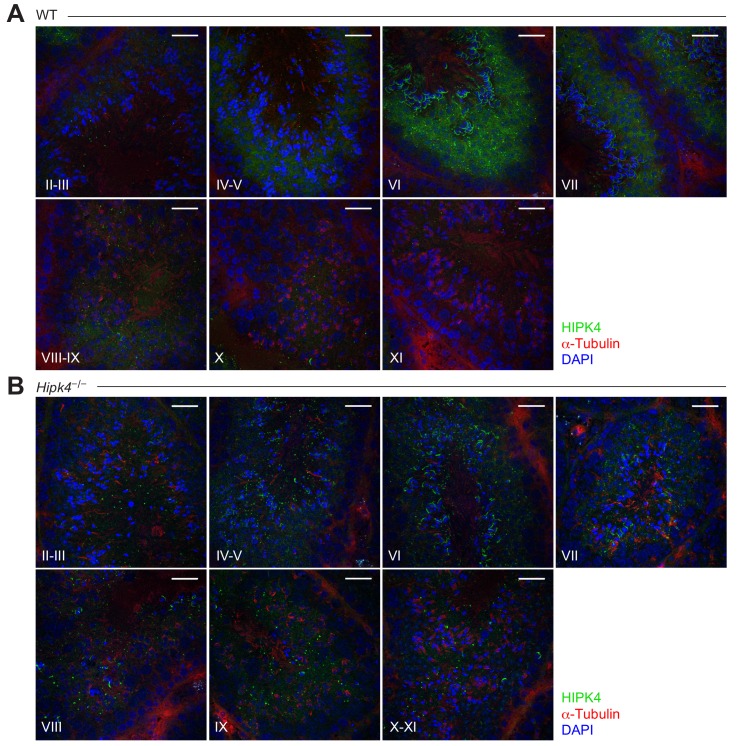
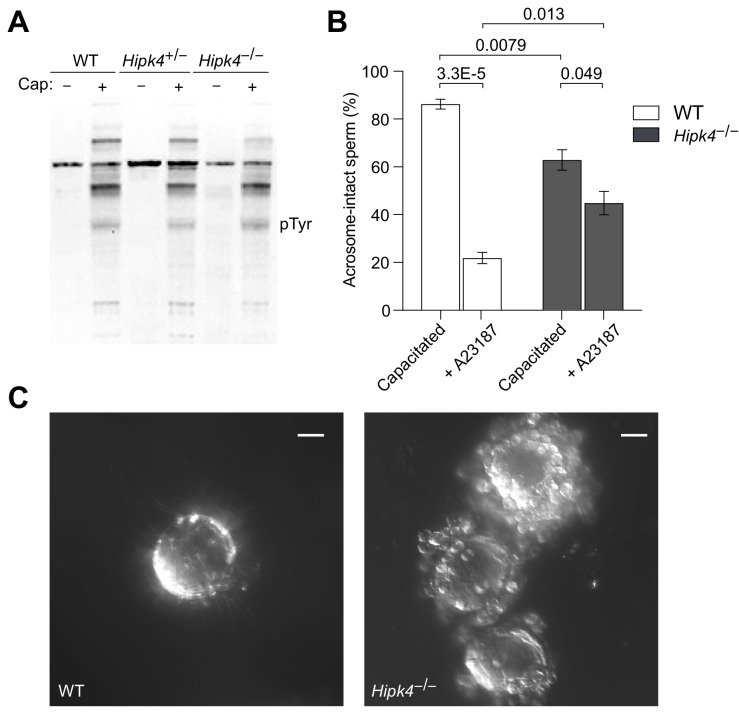
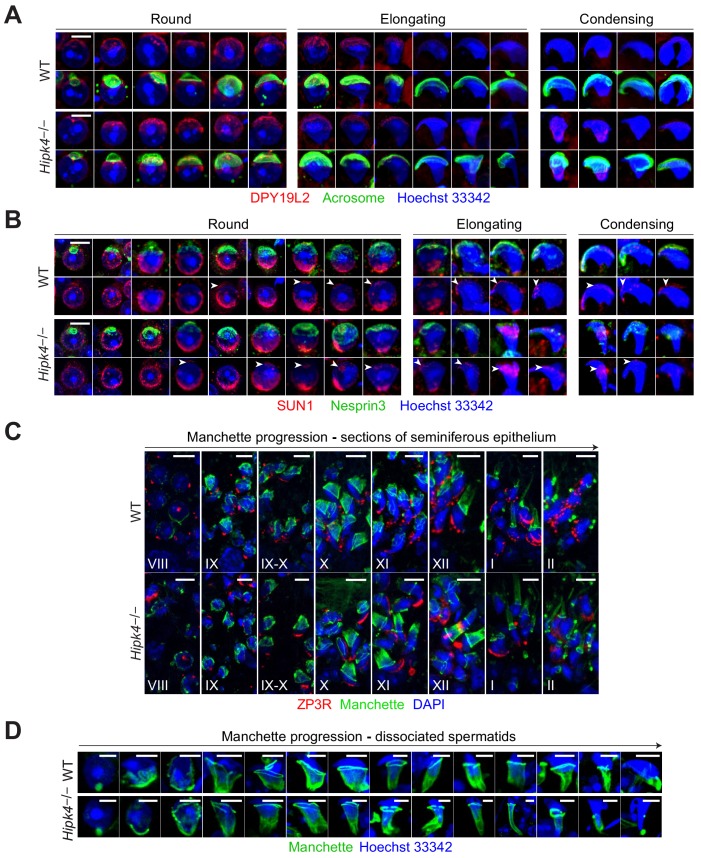
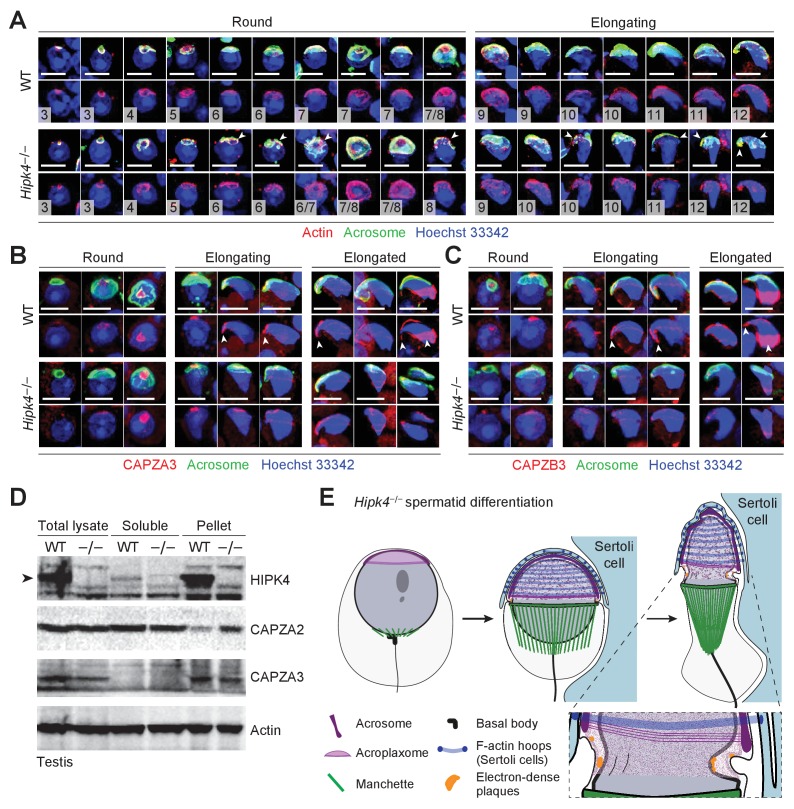
Mammalian spermiogenesis is a remarkable cellular transformation, during which round spermatids elongate into chromatin-condensed spermatozoa. The signaling pathways that coordinate this process are not well understood, and we demonstrate here that homeodomain-interacting protein kinase 4 (HIPK4) is essential for spermiogenesis and male fertility in mice. HIPK4 is predominantly expressed in round and early elongating spermatids, and knockout males are sterile, exhibiting phenotypes consistent with oligoasthenoteratozoospermia. mutant sperm have reduced oocyte binding and are incompetent for in vitro fertilization, but they can still produce viable offspring via intracytoplasmic sperm injection. Optical and electron microscopy of HIPK4-null male germ cells reveals defects in the filamentous actin (F-actin)-scaffolded acroplaxome during spermatid elongation and abnormal head morphologies in mature spermatozoa. We further observe that HIPK4 overexpression induces branched F-actin structures in cultured fibroblasts and that HIPK4 deficiency alters the subcellular distribution of an F-actin capping protein in the testis, supporting a role for this kinase in cytoskeleton remodeling. Our findings establish HIPK4 as an essential regulator of sperm head shaping and potential target for male contraception.
哺乳动物精子发生是一个显著的细胞转化过程,在此过程中,圆形精子细胞伸长为染色质浓缩的精子。协调这一过程的信号通路尚未得到很好的理解,我们在这里证明,同源结构域相互作用蛋白激酶 4(HIPK4)对于小鼠精子发生和雄性生育力是必不可少的。HIPK4主要在圆形和早期伸长的精子细胞中表达,而 HIPK4 敲除雄性是不育的,表现出与少精症、弱精症和畸形精子症一致的表型。突变精子的卵母细胞结合能力降低,体外受精能力不足,但仍可通过胞质内精子注射产生有活力的后代。HIPK4 缺失雄性生殖细胞的光镜和电镜观察显示,在精子细胞伸长过程中丝状肌动蛋白(F-actin)支架的顶体板出现缺陷,成熟精子头部形态异常。我们进一步观察到 HIPK4 过表达在培养的成纤维细胞中诱导分支 F-actin 结构,而 HIPK4 缺失改变了睾丸中 F-actin 盖帽蛋白的亚细胞分布,支持该激酶在细胞骨架重塑中的作用。我们的研究结果确立了 HIPK4 作为精子头部成形的必需调节剂和男性避孕的潜在靶点。