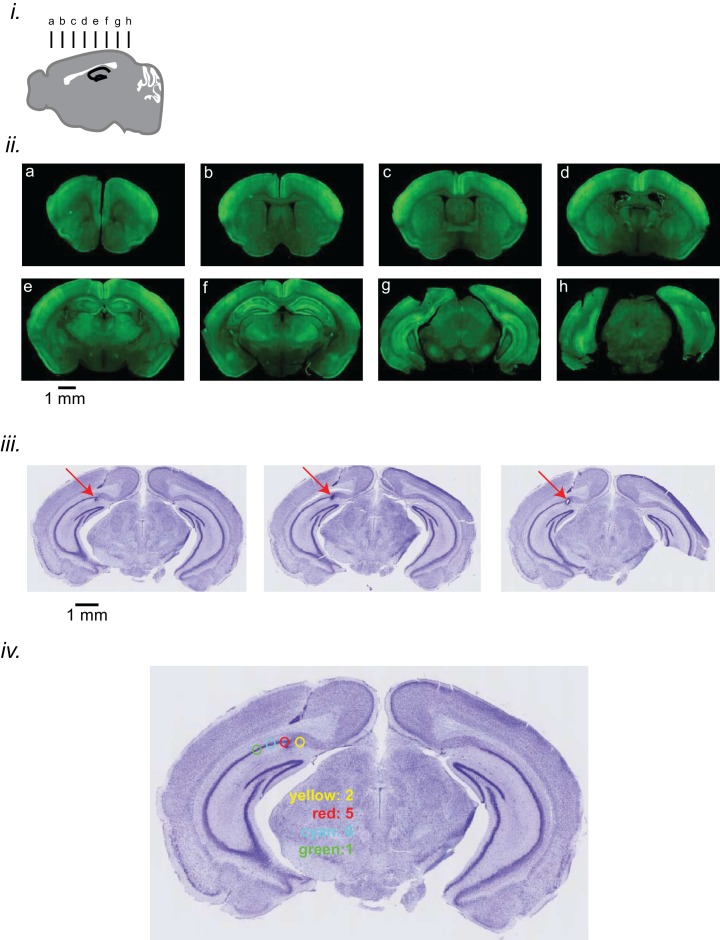
Canadian Centre for Behavioral Neuroscience, University of Lethbridge, Lethbridge, Canada.
Department of Electrical Engineering and Computer Science, University of California, Irvine, United States.
Elife. 2020 Mar 13;9:e51972. doi: 10.7554/eLife.51972.
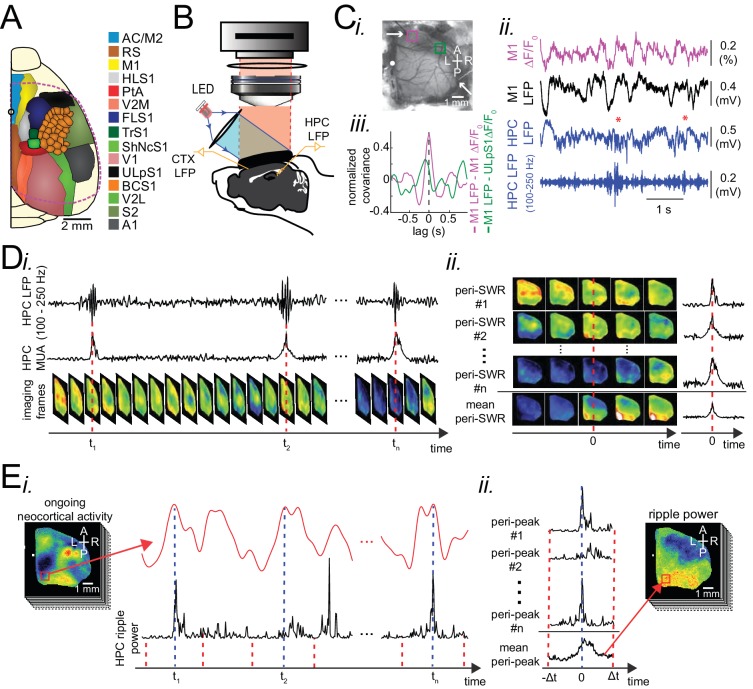
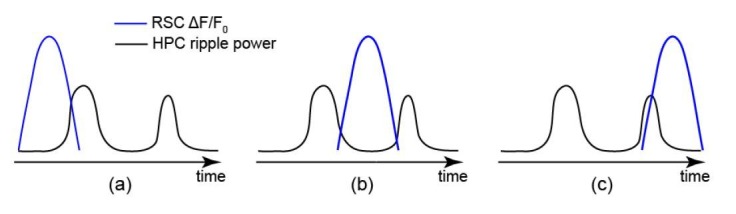
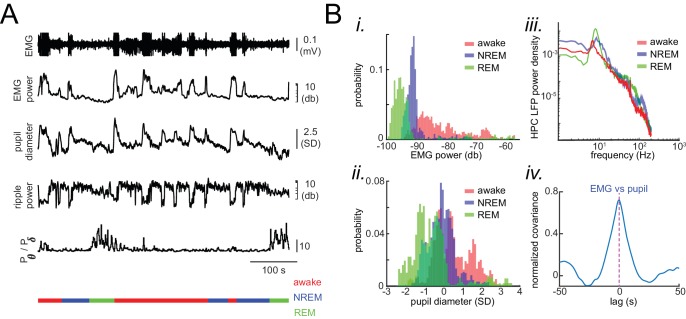
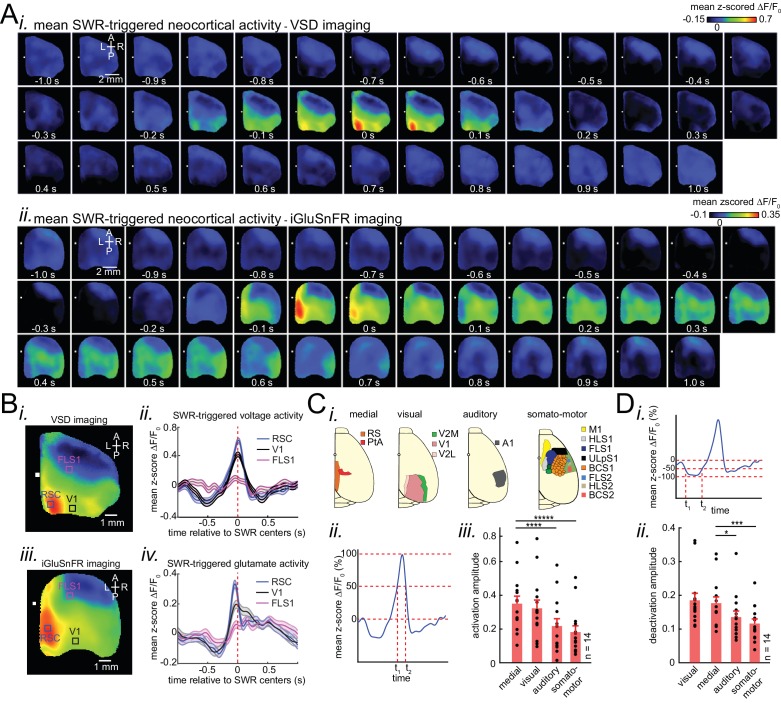
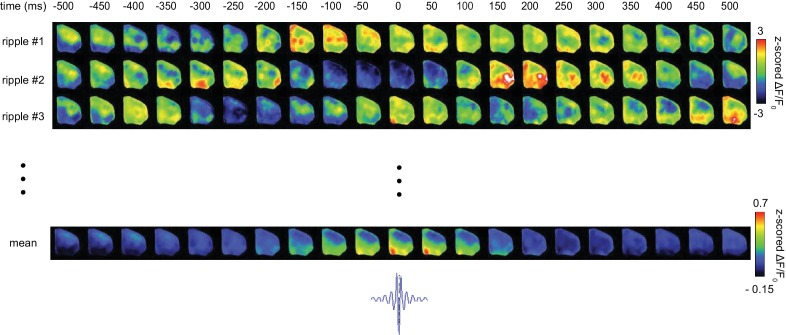
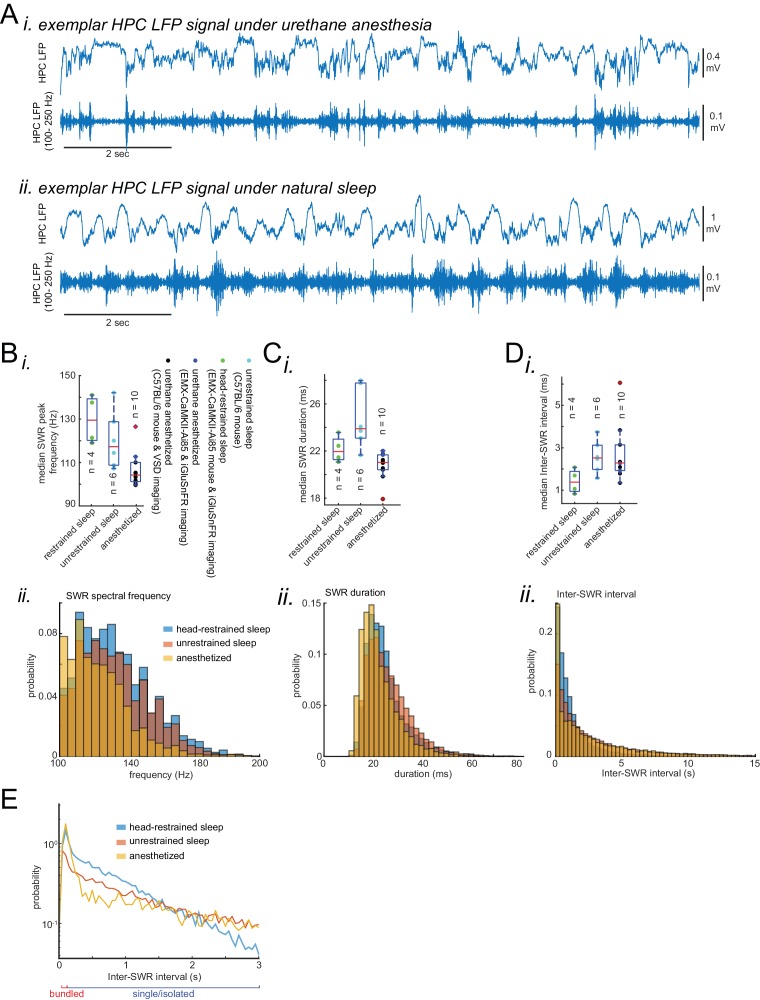
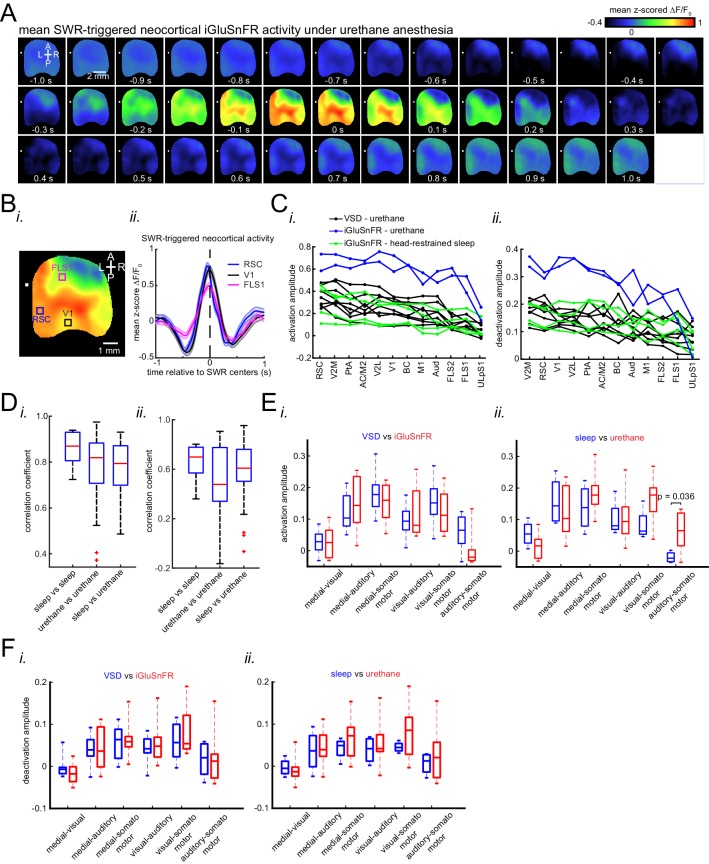
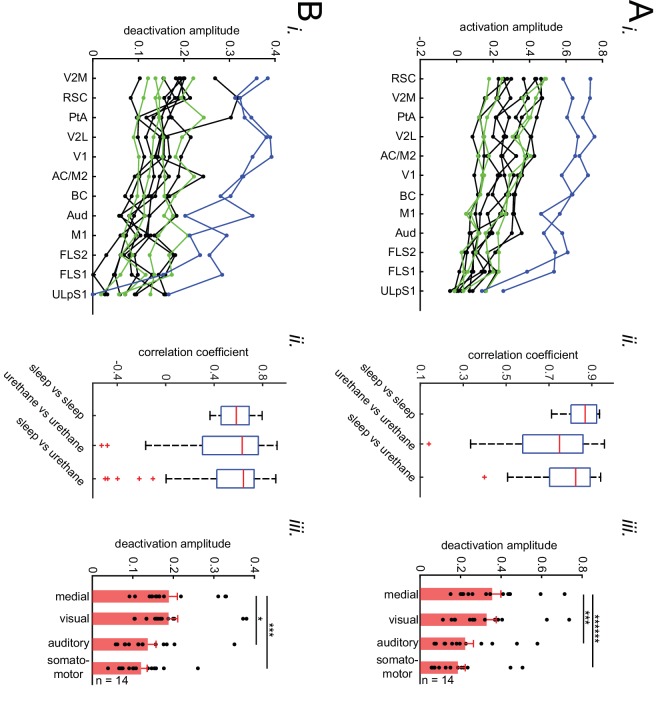
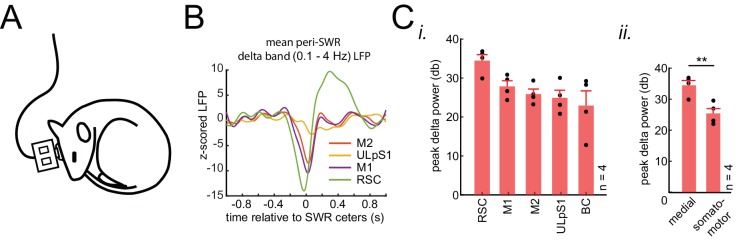
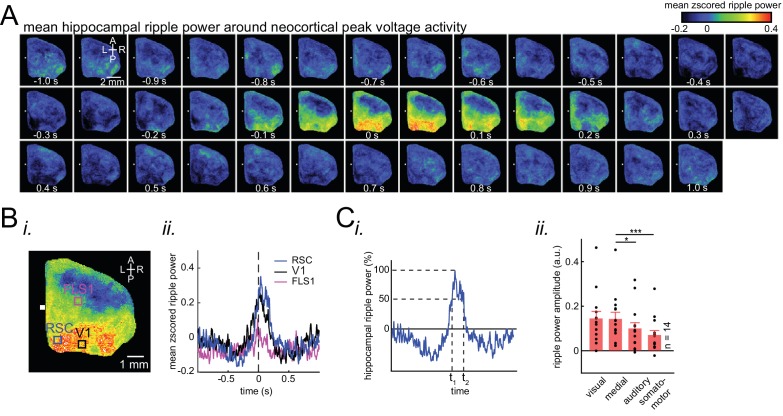
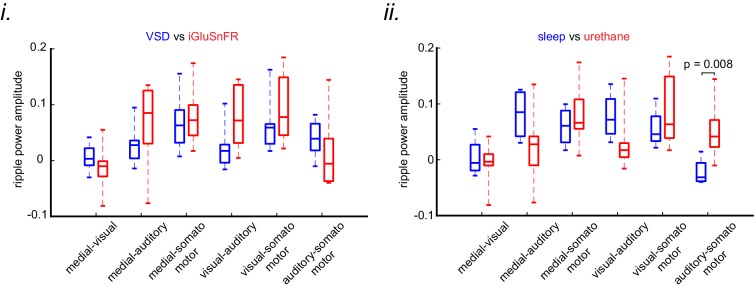
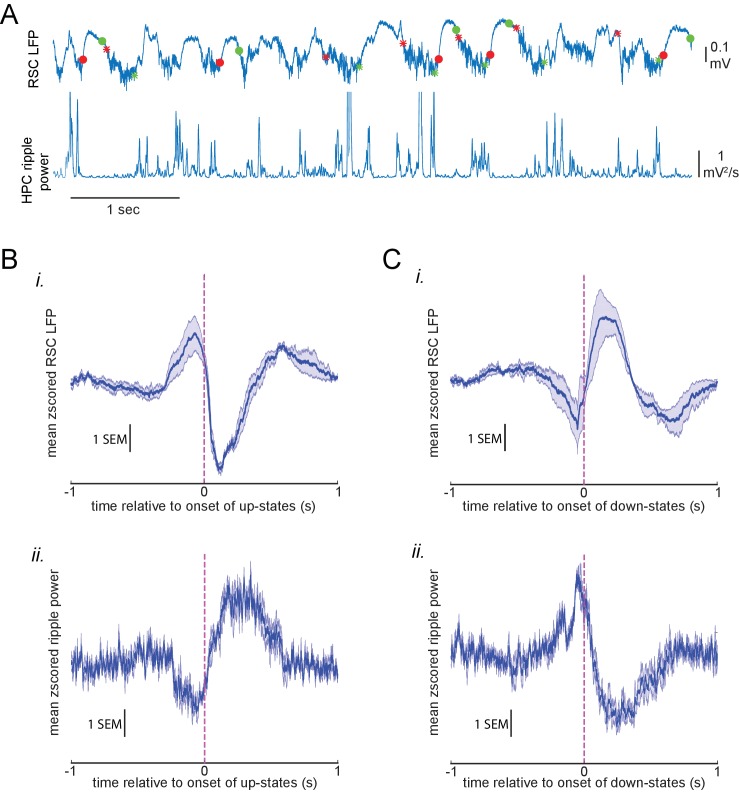
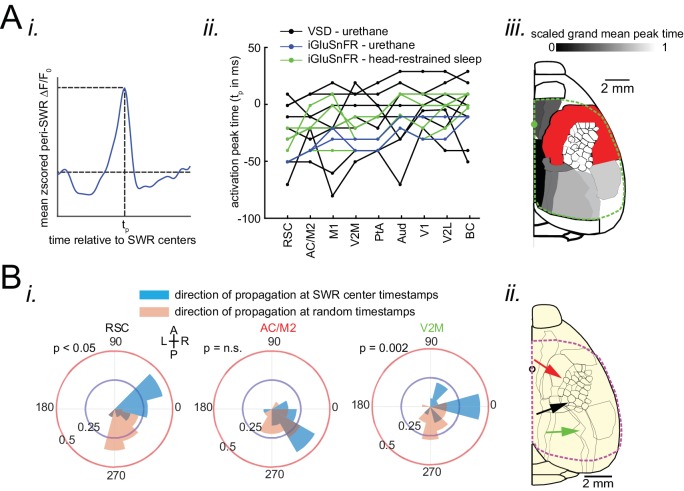
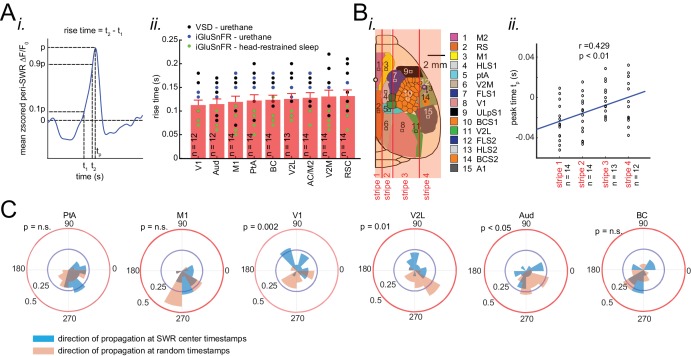
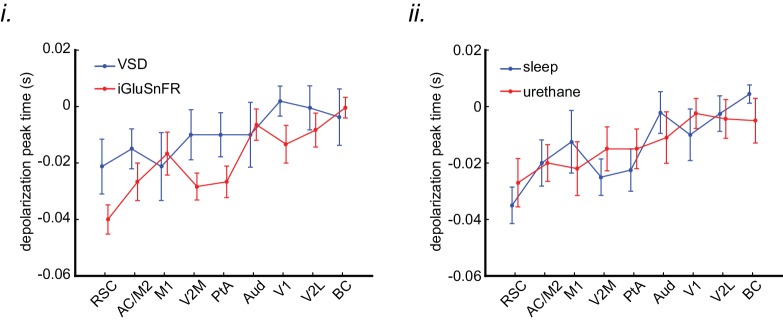
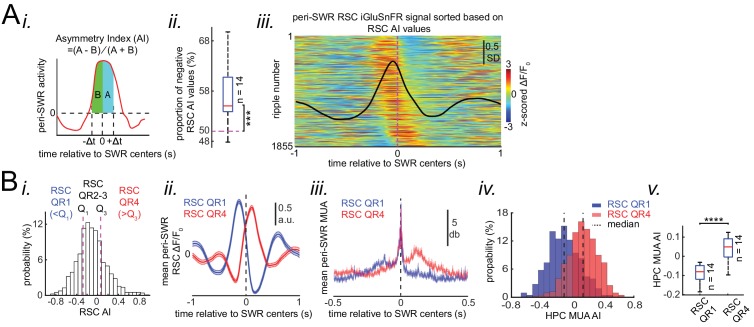
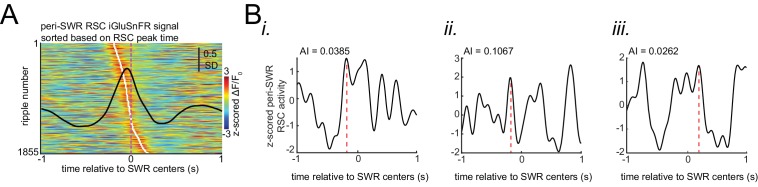
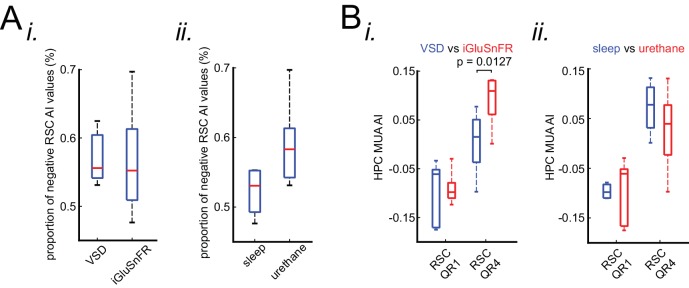
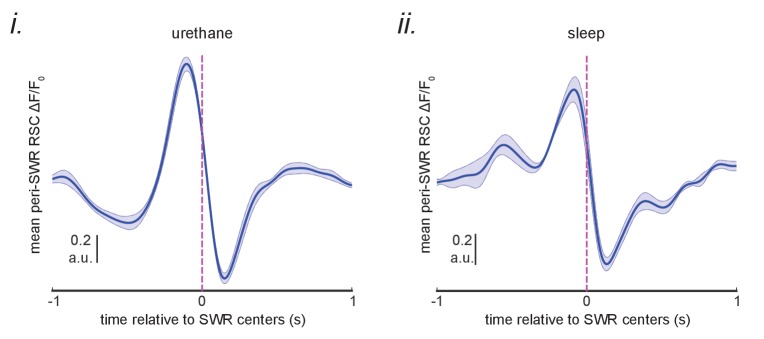
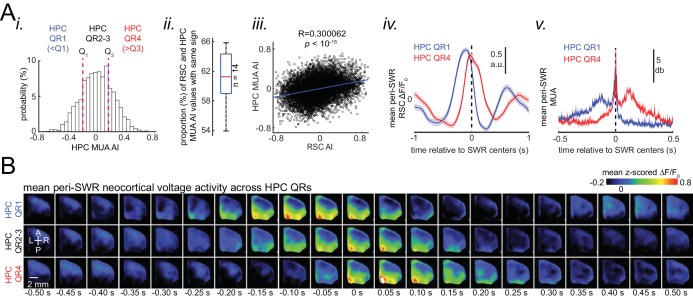
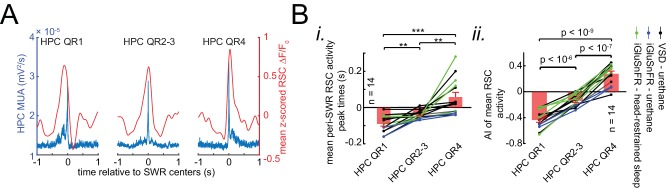
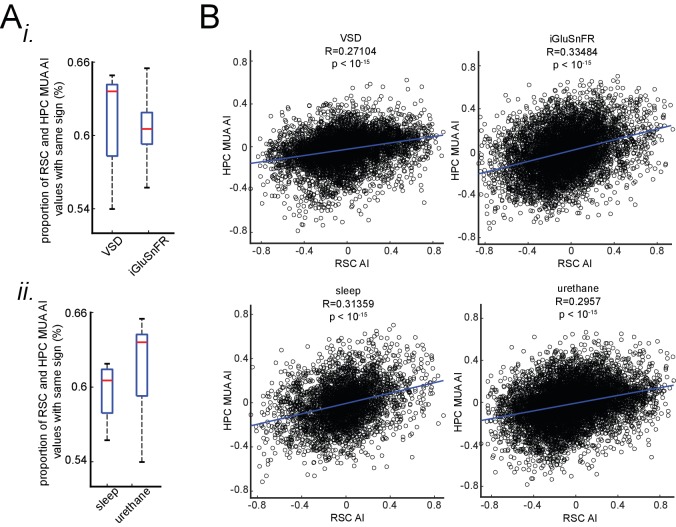
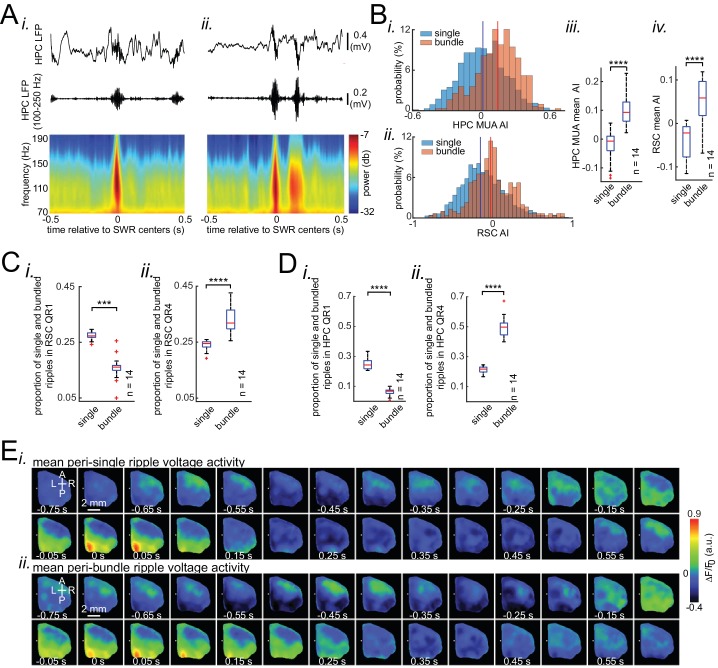
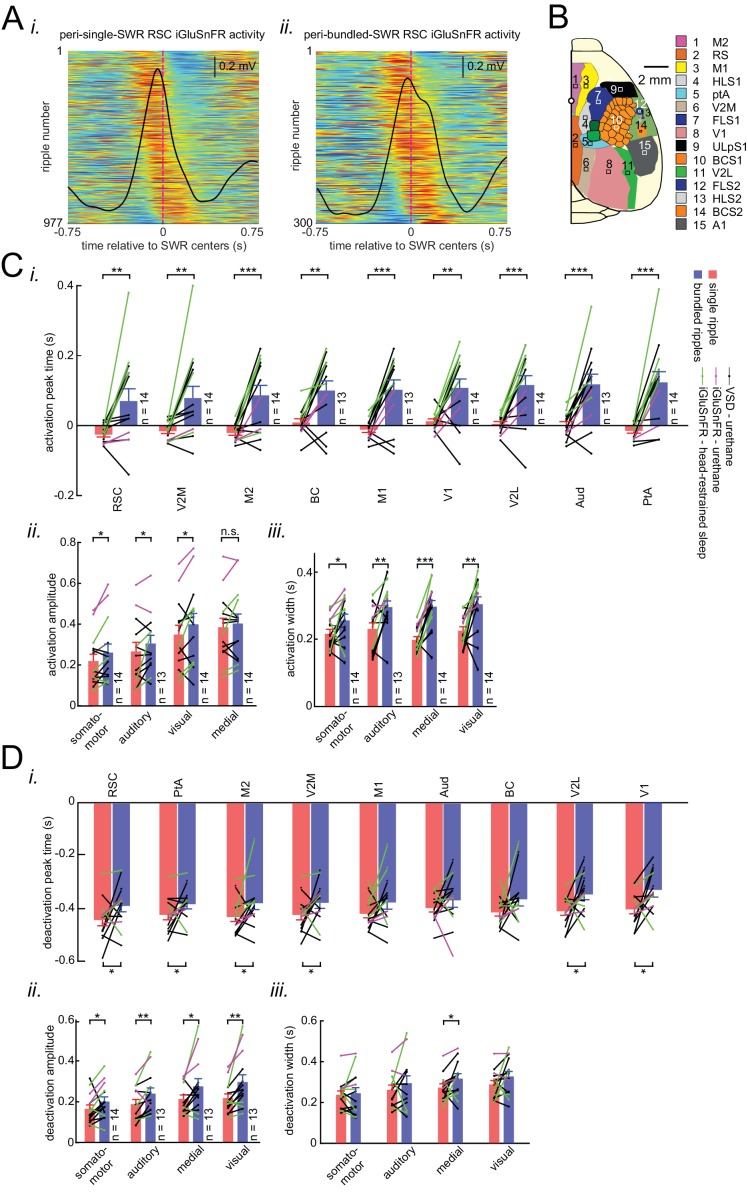
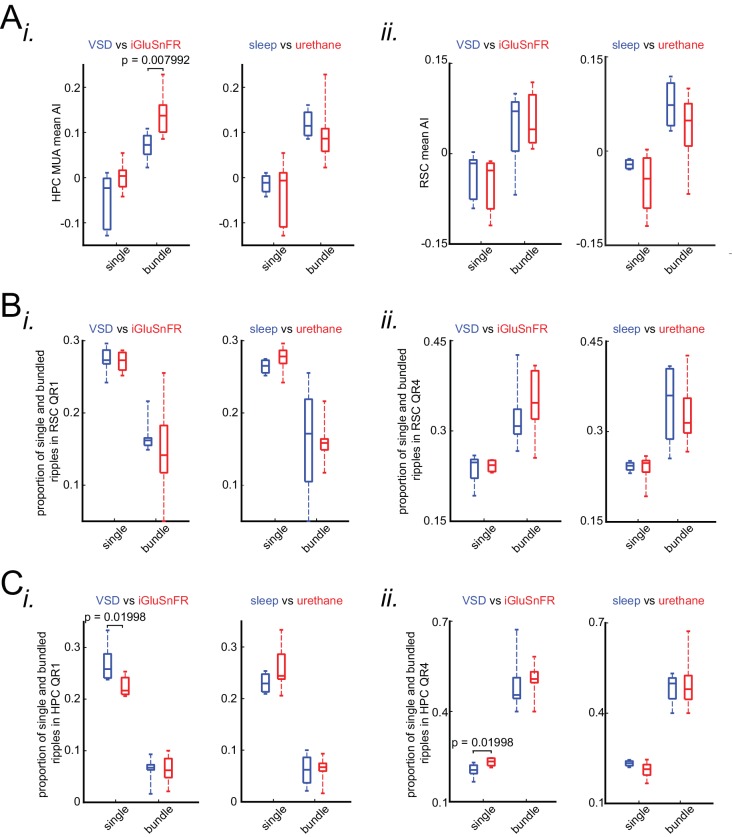
A prevalent model is that sharp-wave ripples (SWR) arise 'spontaneously' in CA3 and propagate recent memory traces outward to the neocortex to facilitate memory consolidation there. Using voltage and extracellular glutamate transient recording over widespread regions of mice dorsal neocortex in relation to CA1 multiunit activity (MUA) and SWR, we find that the largest SWR-related modulation occurs in retrosplenial cortex; however, contrary to the unidirectional hypothesis, neocortical activation exhibited a continuum of activation timings relative to SWRs, varying from leading to lagging. Thus, contrary to the model in which SWRs arise 'spontaneously' in the hippocampus, neocortical activation often precedes SWRs and may thus constitute a trigger event in which neocortical information seeds associative reactivation of hippocampal 'indices'. This timing continuum is consistent with a dynamics in which older, more consolidated memories may in fact initiate the hippocampal-neocortical dialog, whereas reactivation of newer memories may be initiated predominantly in the hippocampus.
一种流行的模式是,尖波涟漪(SWR)“自发”出现在 CA3 中,并将最近的记忆痕迹向外传播到新皮层,以促进那里的记忆巩固。使用电压和细胞外谷氨酸瞬变记录与 CA1 多单位活动(MUA)和 SWR 相关的小鼠背侧新皮层的广泛区域,我们发现与 SWR 相关的最大调制发生在后穹窿皮质;然而,与单向假说相反,新皮层的激活表现出与 SWR 相关的激活时间的连续体,从领先到滞后不等。因此,与 SWR 在海马体中“自发”产生的模型相反,新皮层的激活通常先于 SWR,因此可能构成一个触发事件,其中新皮层信息引发海马体“指标”的联想再激活。这种时间连续体与一种动力学一致,即较旧、更巩固的记忆实际上可能启动海马体-新皮层对话,而较新记忆的再激活可能主要在海马体中启动。