Department of Biological Sciences, Korea Advanced Institute of Science and Technology, Daejeon 34141, South Korea.
Biology Department, School of Science, Indiana University-Purdue University Indianapolis, Indianapolis, IN 46202, USA.
Nucleic Acids Res. 2020 Jun 4;48(10):5442-5456. doi: 10.1093/nar/gkaa317.
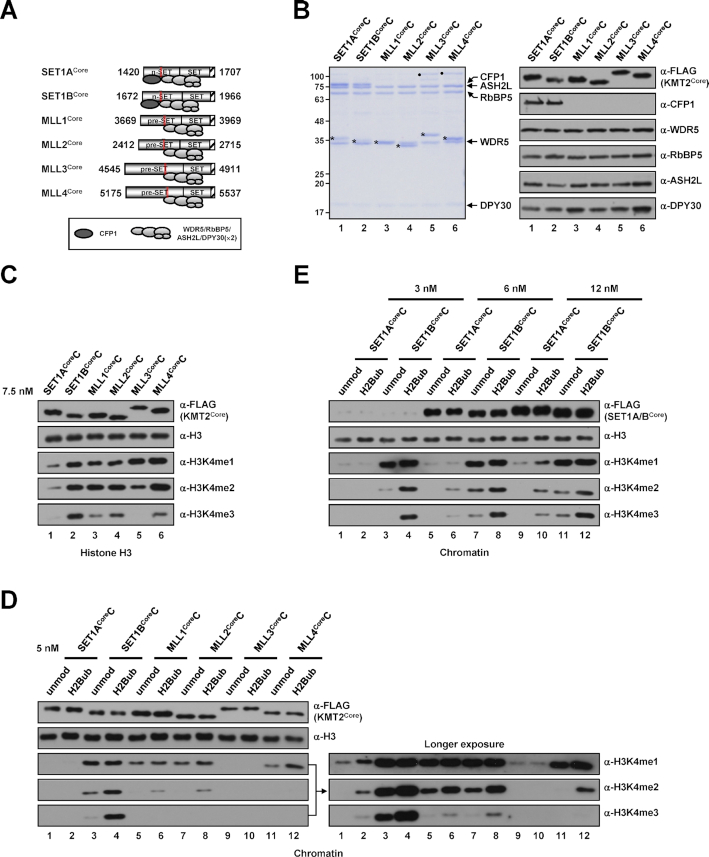
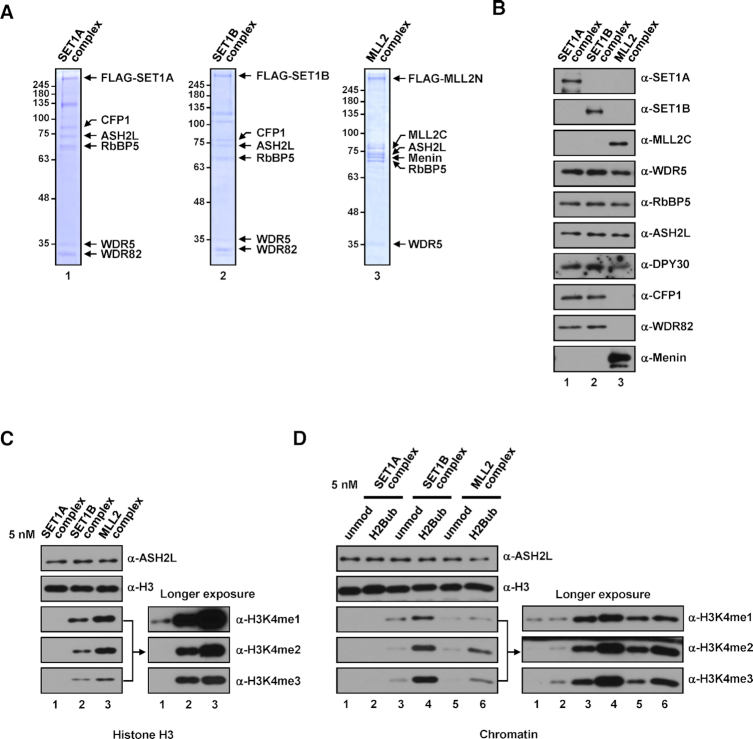
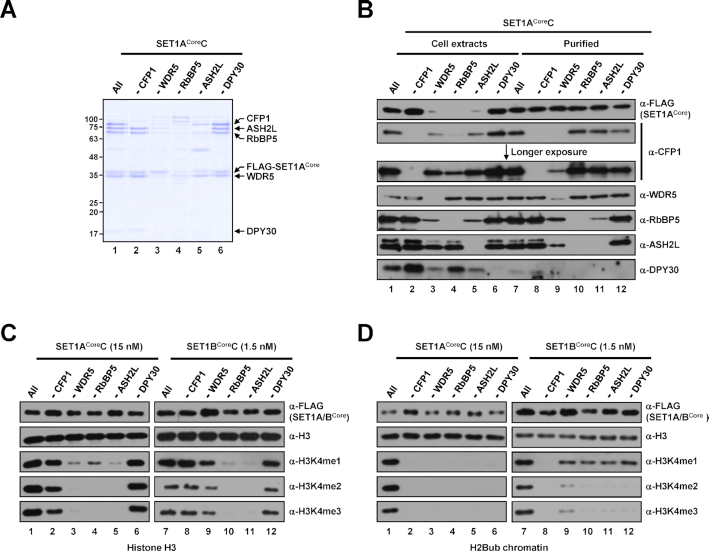
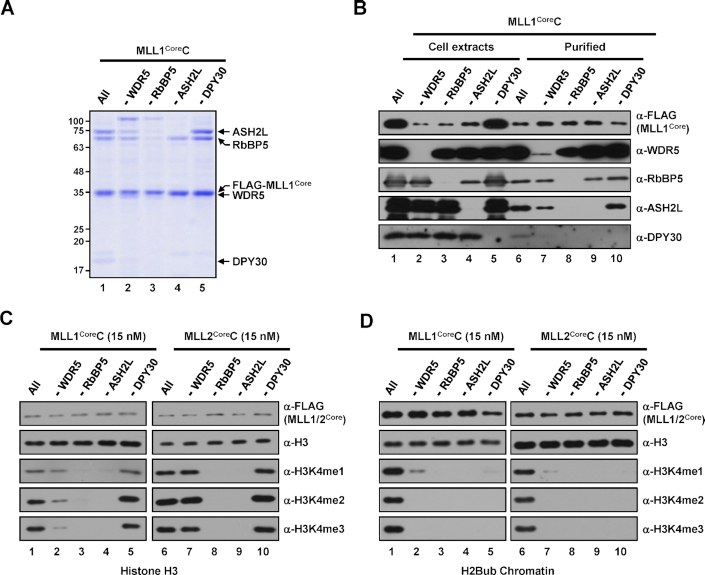
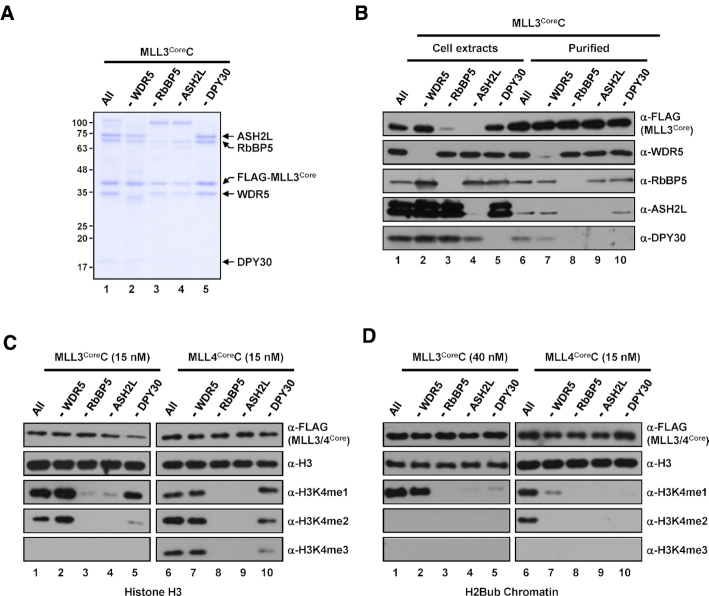
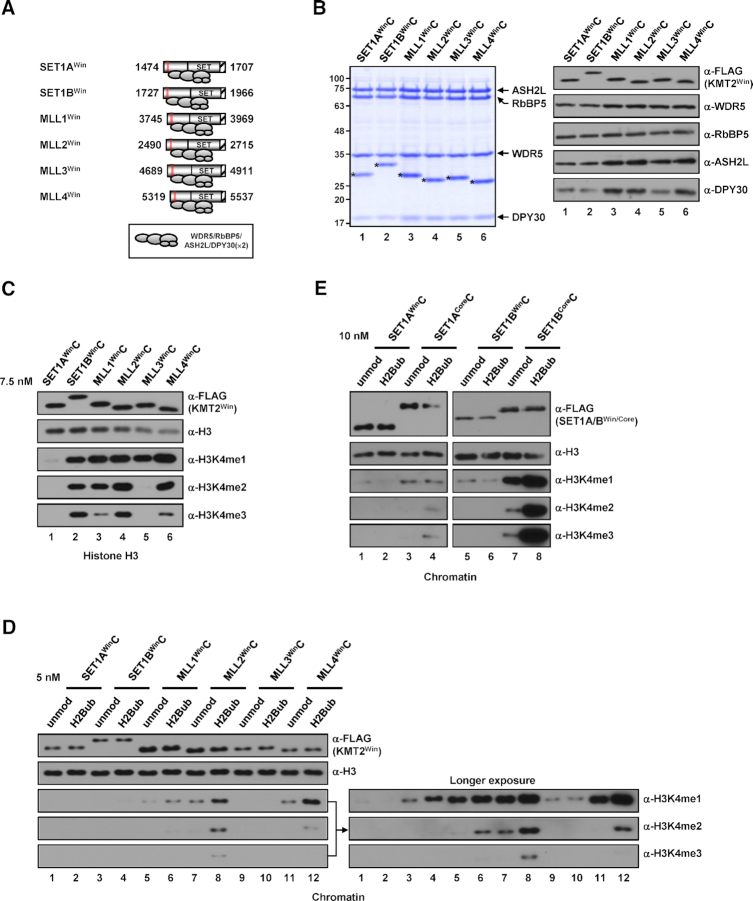
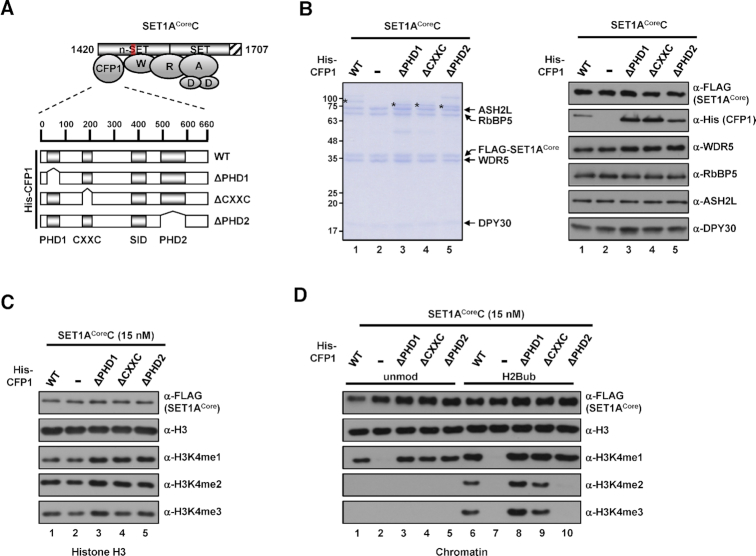
In mammalian cells, distinct H3K4 methylation states are created by deposition of methyl groups by multiple complexes of histone lysine methyltransferase 2 (KMT2) family proteins. For comprehensive analyses that directly compare the catalytic properties of all six human KMT2 complexes, we employed a biochemically defined system reconstituted with recombinant KMT2 core complexes (KMT2CoreCs) containing minimal components required for nucleosomal H3K4 methylation activity. We found that each KMT2CoreC generates distinct states and different levels of H3K4 methylation, and except for MLL3 all are stimulated by H2Bub. Notably, SET1BCoreC exhibited the strongest H3K4 methylation activity and, to our surprise, did not require H2B ubiquitylation (H2Bub); in contrast, H2Bub was required for the H3K4me2/3 activity of the paralog SET1ACoreC. We also found that WDR5, RbBP5, ASH2L and DPY30 are required for efficient H3K4 methyltransferase activities of all KMT2CoreCs except MLL3, which could produce H3K4me1 in the absence of WDR5. Importantly, deletion of the PHD2 domain of CFP1 led to complete loss of the H3K4me2/3 activities of SET1A/BCoreCs in the presence of H2Bub, indicating a critical role for this domain in the H2Bub-stimulated H3K4 methylation. Collectively, our results suggest that each KMT2 complex methylates H3K4 through distinct mechanisms in which individual subunits differentially participate.
在哺乳动物细胞中,不同的 H3K4 甲基化状态是由多个组蛋白赖氨酸甲基转移酶 2(KMT2)家族蛋白复合物沉积甲基基团形成的。为了进行全面分析,直接比较六种人源 KMT2 复合物的催化特性,我们采用了一种生物化学定义的系统,该系统由含有核小体 H3K4 甲基化活性所需最小成分的重组 KMT2 核心复合物(KMT2CoreCs)重建。我们发现每个 KMT2CoreC 产生不同的状态和不同水平的 H3K4 甲基化,除了 MLL3 之外,所有的 KMT2CoreC 都受到 H2Bub 的刺激。值得注意的是,SET1BCoreC 表现出最强的 H3K4 甲基化活性,而且令我们惊讶的是,它不需要 H2B 泛素化(H2Bub);相比之下,H2Bub 是 SET1ACoreC 的 H3K4me2/3 活性所必需的。我们还发现,WDR5、RbBP5、ASH2L 和 DPY30 是除 MLL3 之外所有 KMT2CoreC 的有效 H3K4 甲基转移酶活性所必需的,MLL3 可以在没有 WDR5 的情况下产生 H3K4me1。重要的是,CFP1 的 PHD2 结构域缺失导致 SET1A/BCoreCs 在 H2Bub 存在的情况下完全丧失 H3K4me2/3 活性,表明该结构域在 H2Bub 刺激的 H3K4 甲基化中起着关键作用。总之,我们的结果表明,每个 KMT2 复合物通过不同的机制甲基化 H3K4,其中各个亚基以不同的方式参与。