Department of Pharmacology, University of California San Diego, La Jolla, United States.
Department of Pharmacology and Molecular Sciences, The Johns Hopkins University School of Medicine, Baltimore, United States.
Elife. 2020 May 26;9:e57410. doi: 10.7554/eLife.57410.
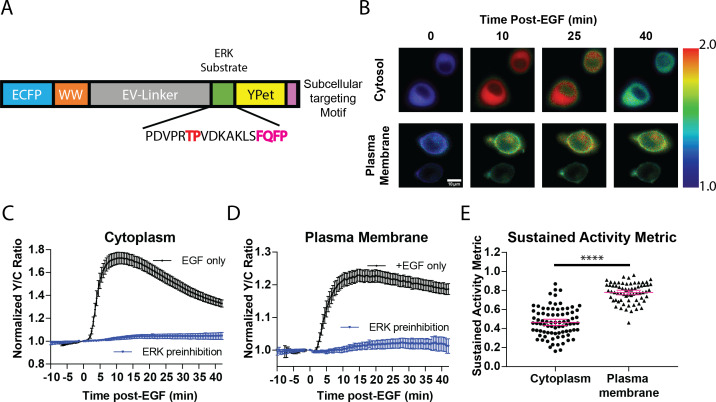
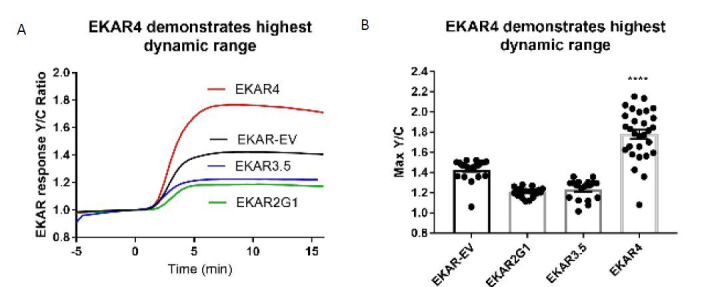
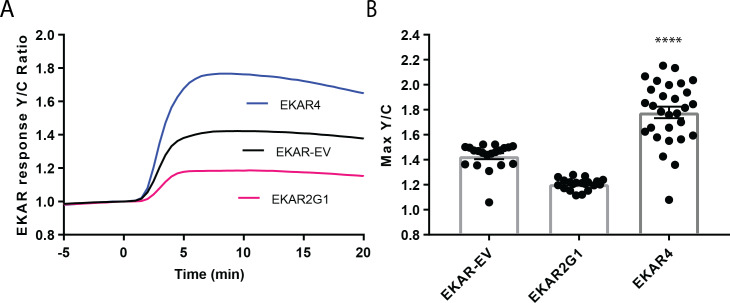
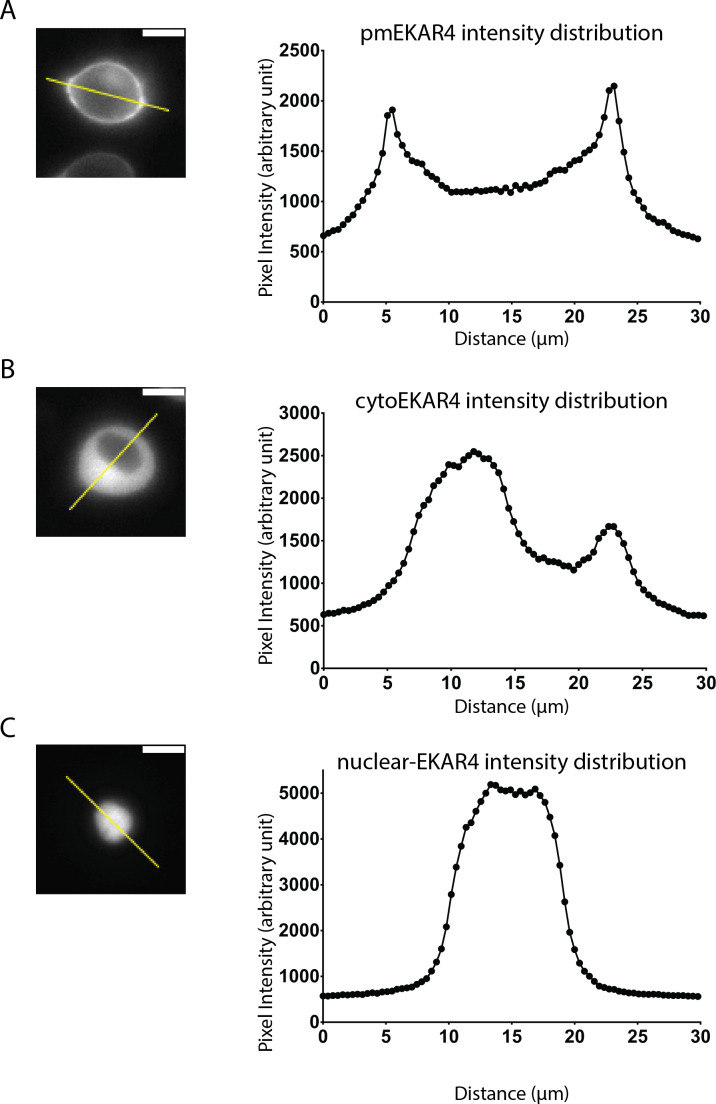
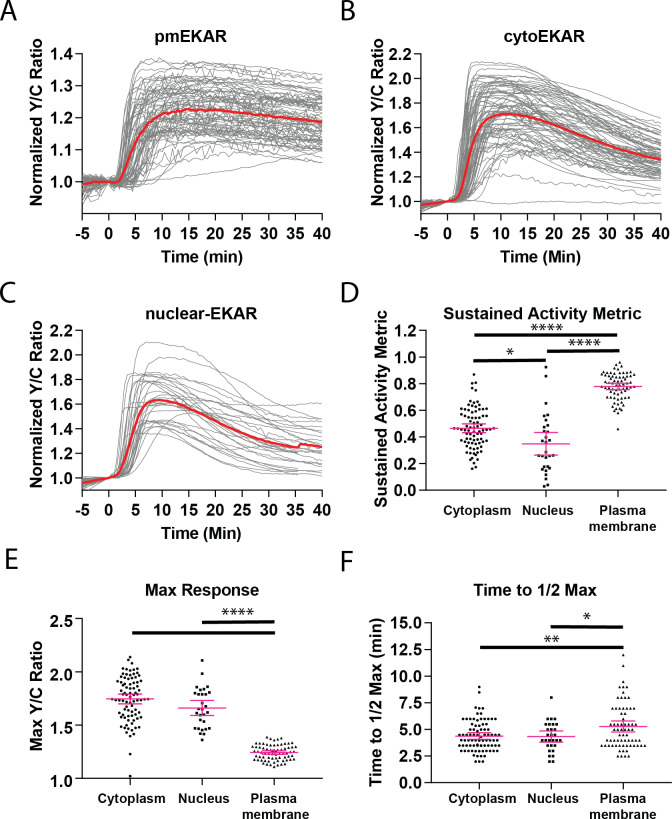
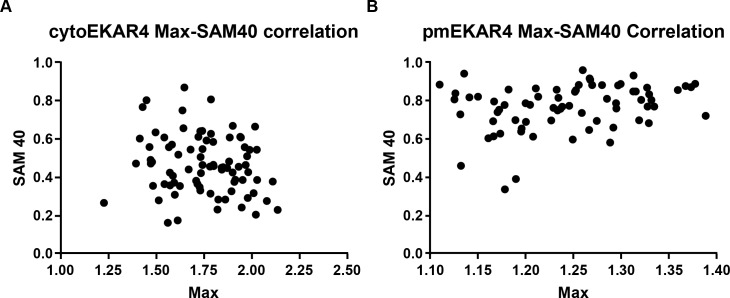
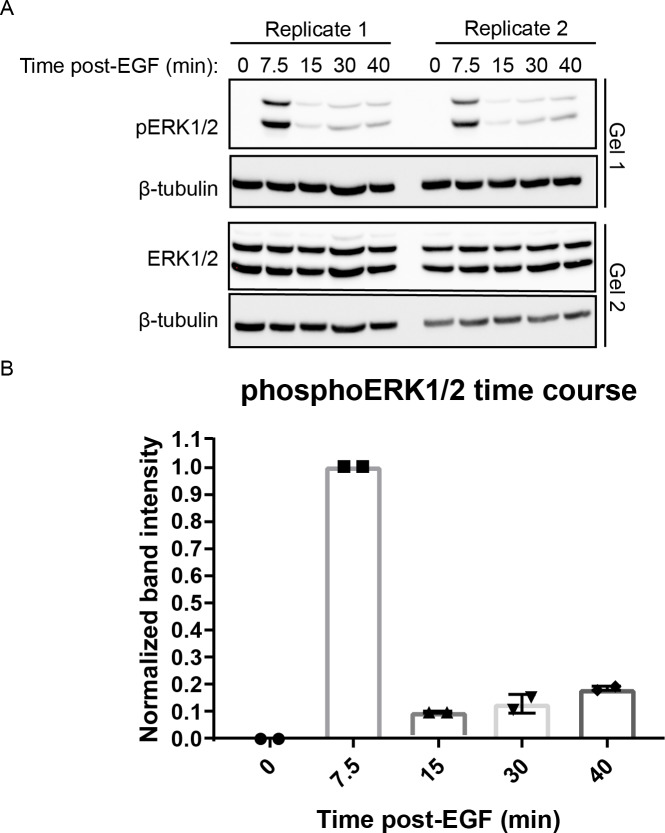
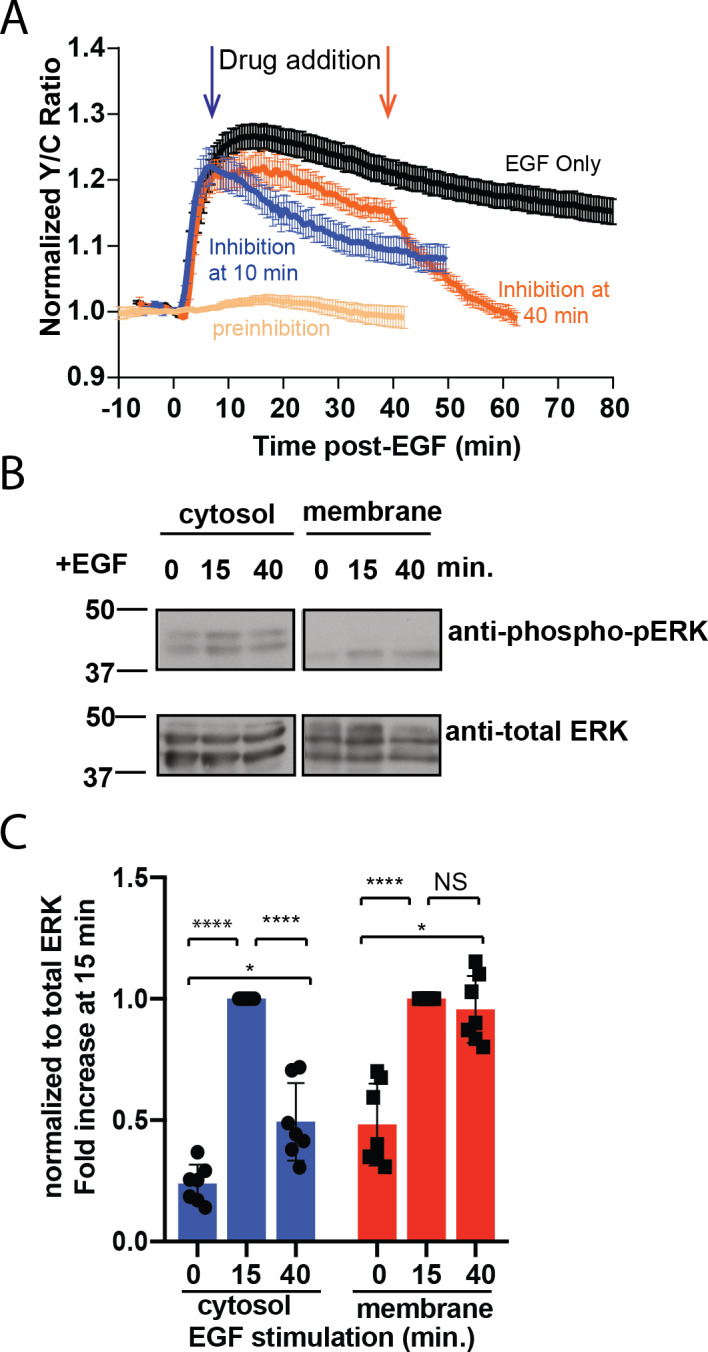
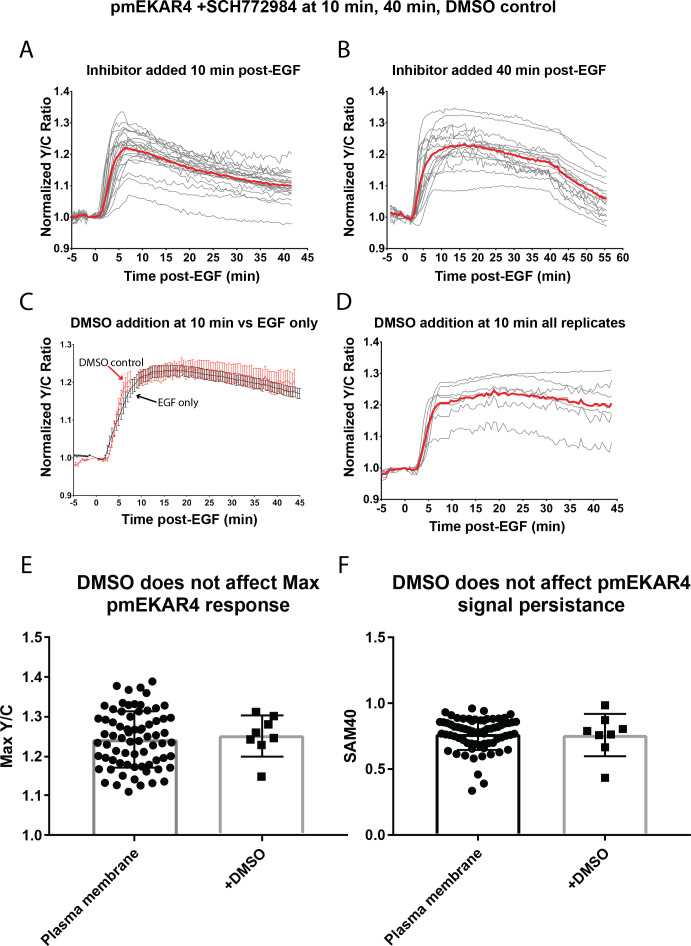
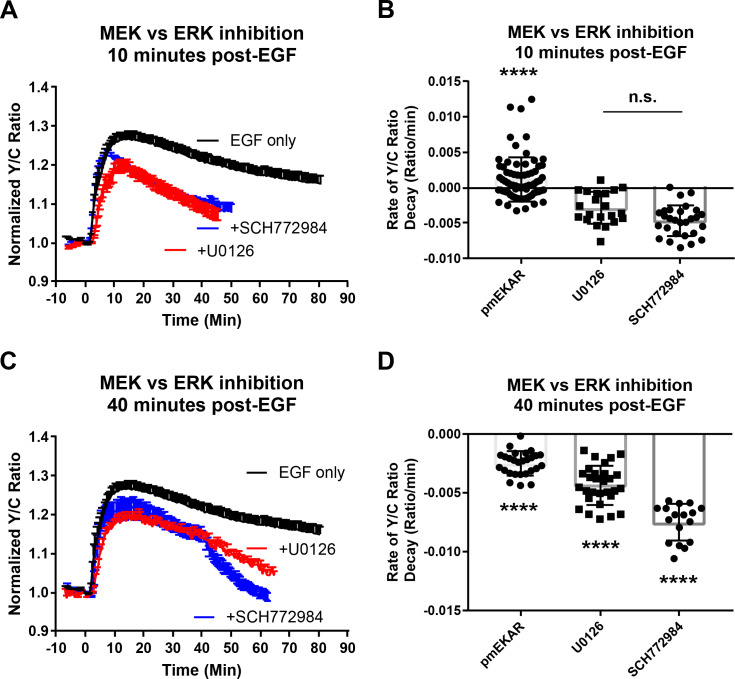
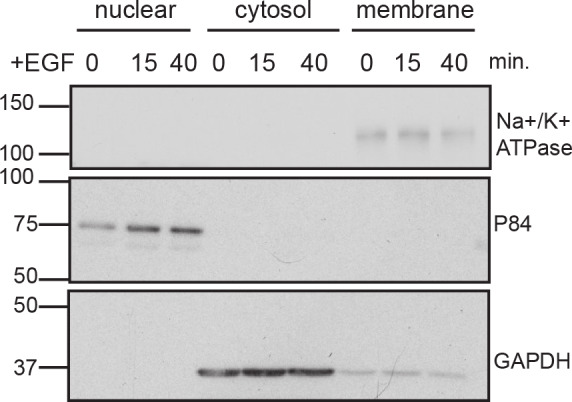
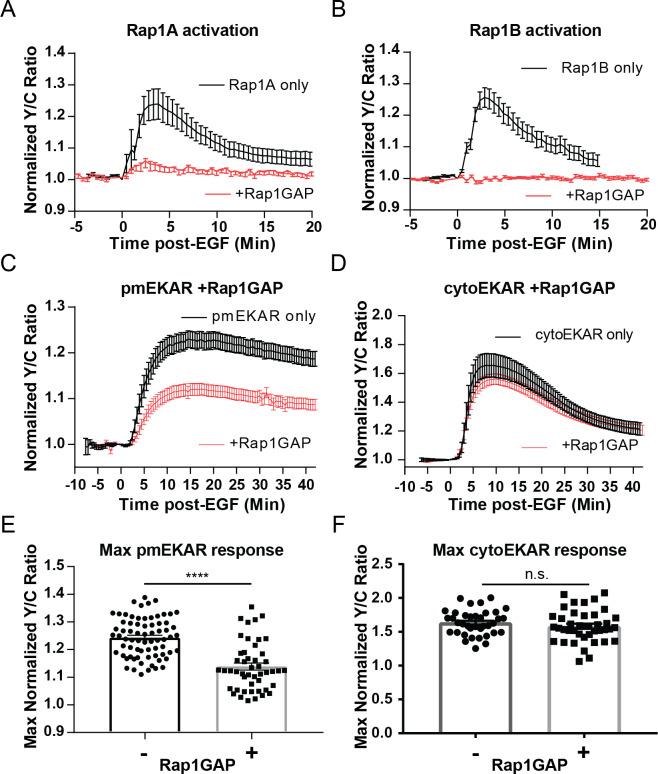
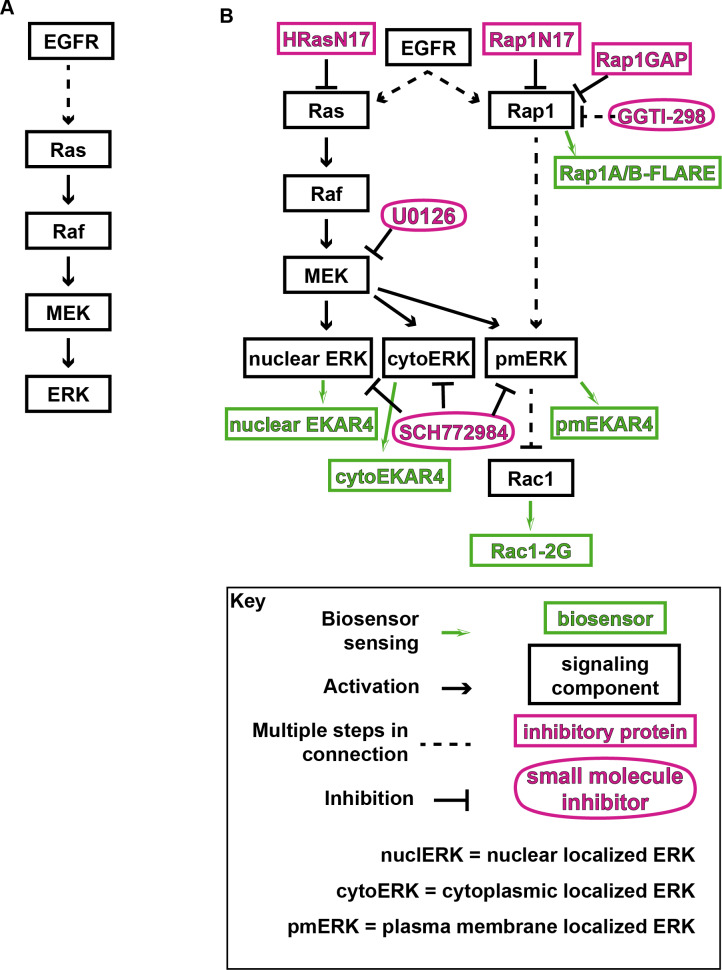
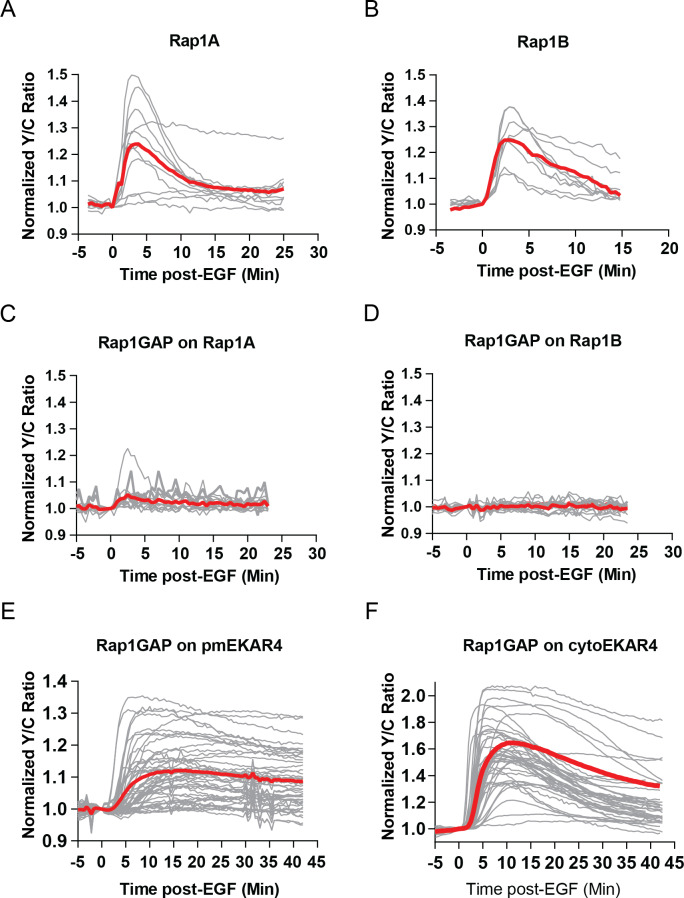
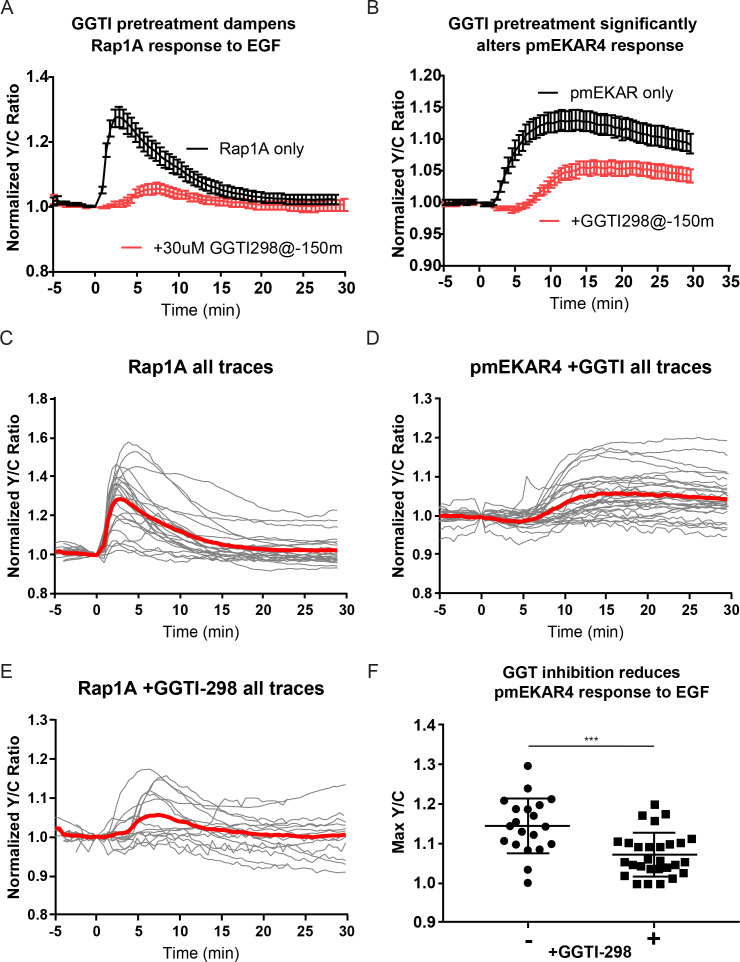
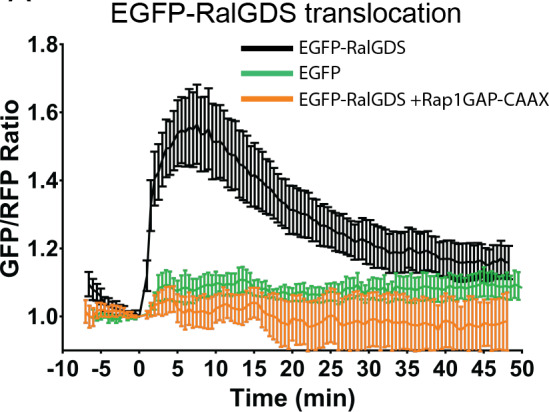
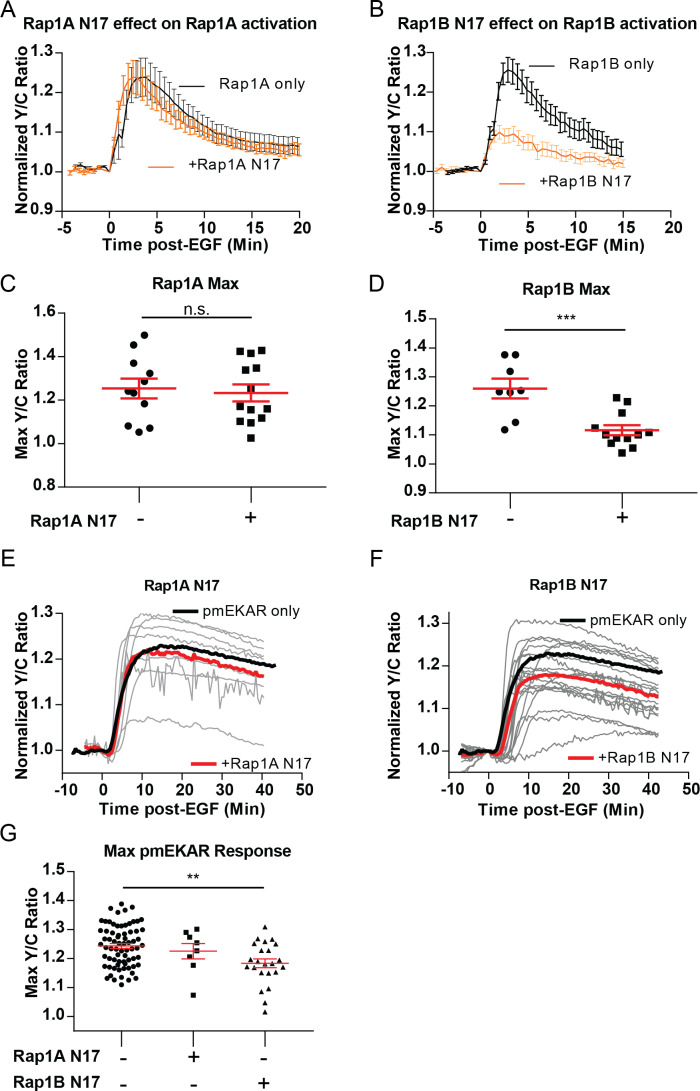
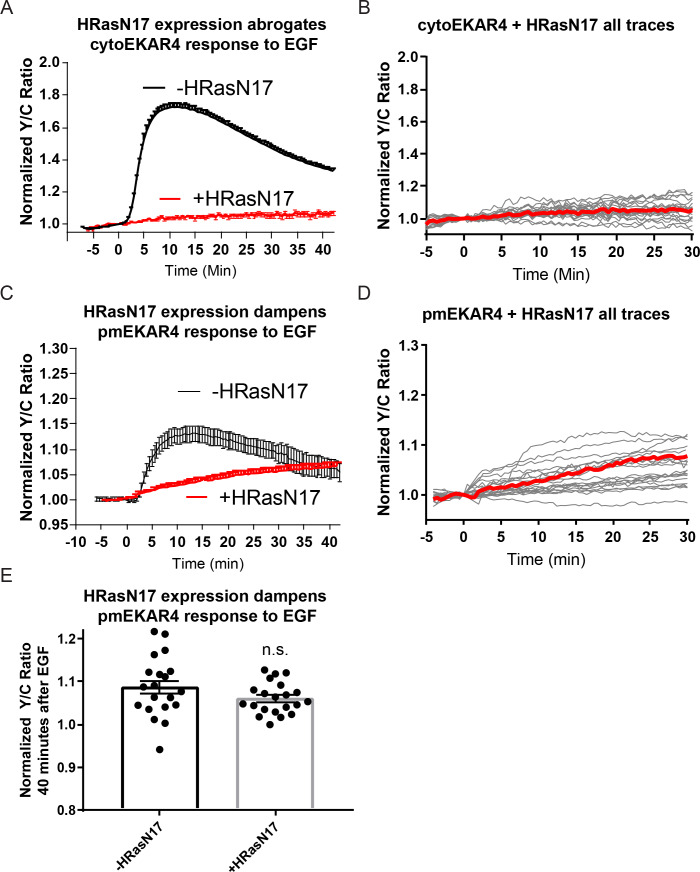
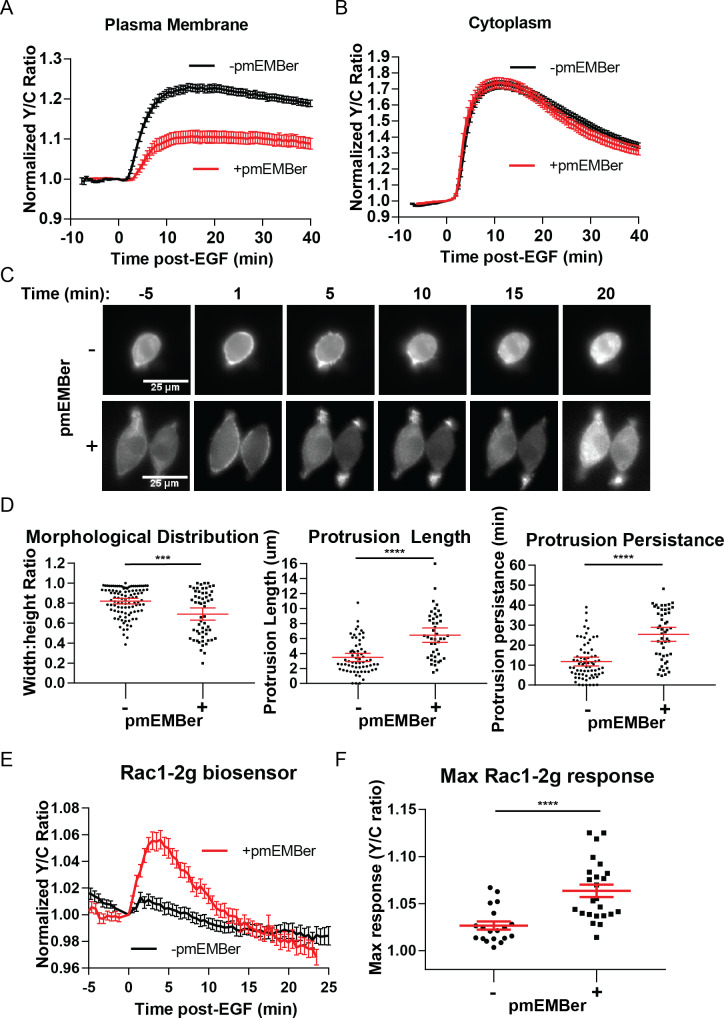
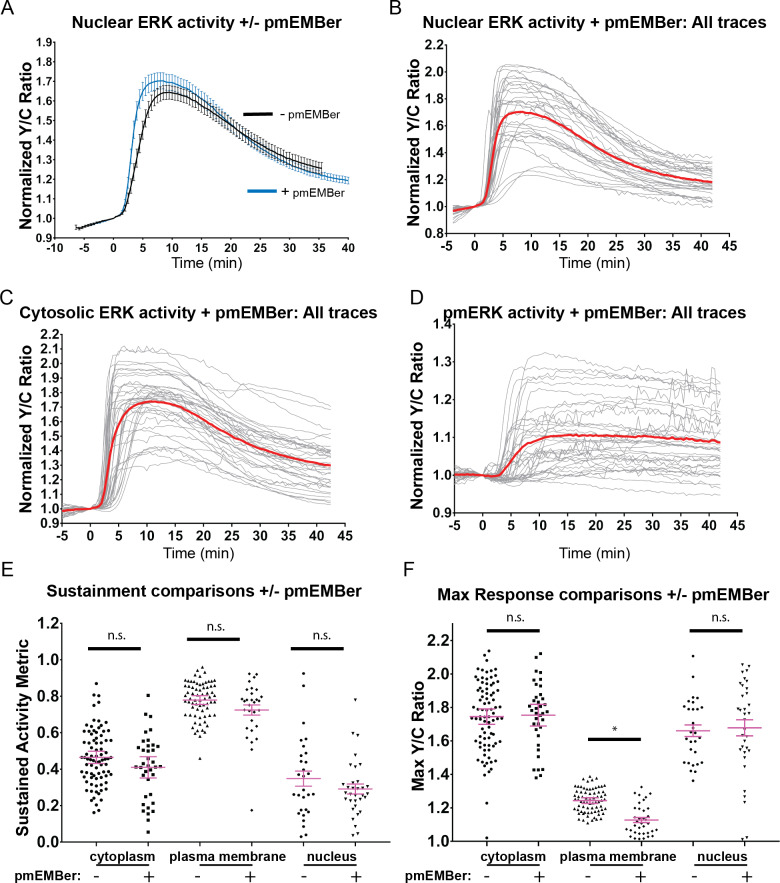
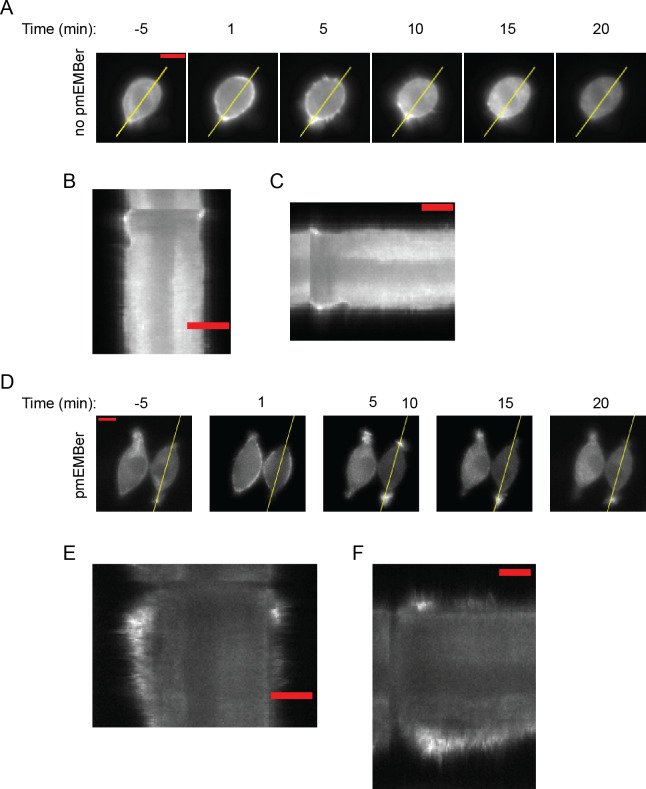
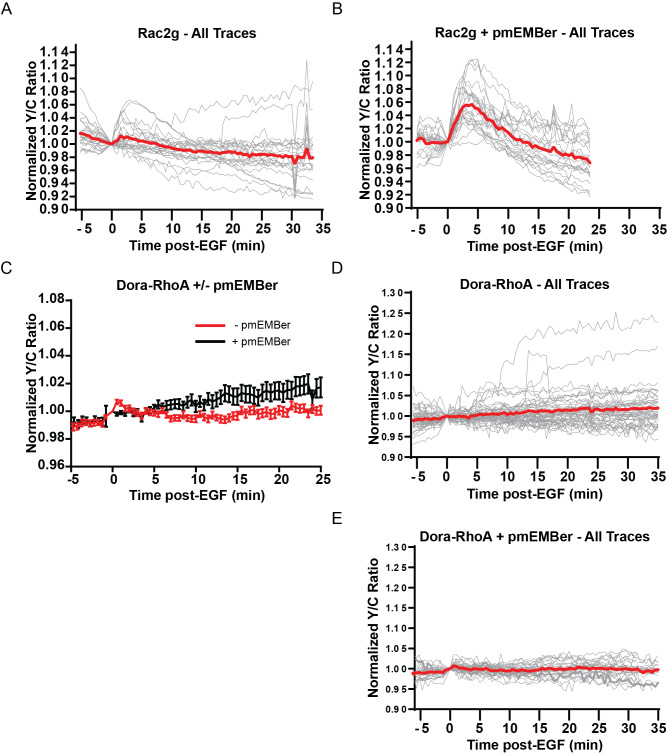
A variety of different signals induce specific responses through a common, extracellular-signal regulated kinase (ERK)-dependent cascade. It has been suggested that signaling specificity can be achieved through precise temporal regulation of ERK activity. Given the wide distrubtion of ERK susbtrates across different subcellular compartments, it is important to understand how ERK activity is temporally regulated at specific subcellular locations. To address this question, we have expanded the toolbox of Förster Resonance Energy Transfer (FRET)-based ERK biosensors by creating a series of improved biosensors targeted to various subcellular regions via sequence specific motifs to measure spatiotemporal changes in ERK activity. Using these sensors, we showed that EGF induces sustained ERK activity near the plasma membrane in sharp contrast to the transient activity observed in the cytoplasm and nucleus. Furthermore, EGF-induced plasma membrane ERK activity involves Rap1, a noncanonical activator, and controls cell morphology and EGF-induced membrane protrusion dynamics. Our work strongly supports that spatial and temporal regulation of ERK activity is integrated to control signaling specificity from a single extracellular signal to multiple cellular processes.
各种不同的信号通过共同的细胞外信号调节激酶 (ERK) 依赖性级联反应诱导特定的反应。有人提出,通过 ERK 活性的精确时间调节可以实现信号特异性。鉴于 ERK 底物在不同的亚细胞隔室中广泛分布,了解 ERK 活性如何在特定的亚细胞位置进行时间调节非常重要。为了解决这个问题,我们通过使用序列特异性基序来靶向各种亚细胞区域,创建了一系列改进的基于Förster 共振能量转移 (FRET) 的 ERK 生物传感器,从而扩展了 ERK 生物传感器的工具包,以测量 ERK 活性的时空变化。使用这些传感器,我们表明 EGF 在质膜附近诱导持续的 ERK 活性,与细胞质和细胞核中观察到的短暂活性形成鲜明对比。此外,EGF 诱导的质膜 ERK 活性涉及 Rap1,一种非典型激活剂,并且控制细胞形态和 EGF 诱导的膜突起动力学。我们的工作强烈支持 ERK 活性的时空调节是整合的,以控制从单个细胞外信号到多个细胞过程的信号特异性。