Department of Biology, University of Iowa, Iowa City, IA 52242-1324.
Mol Biol Cell. 2020 Aug 1;31(17):1951-1961. doi: 10.1091/mbc.E20-02-0139. Epub 2020 Jun 17.
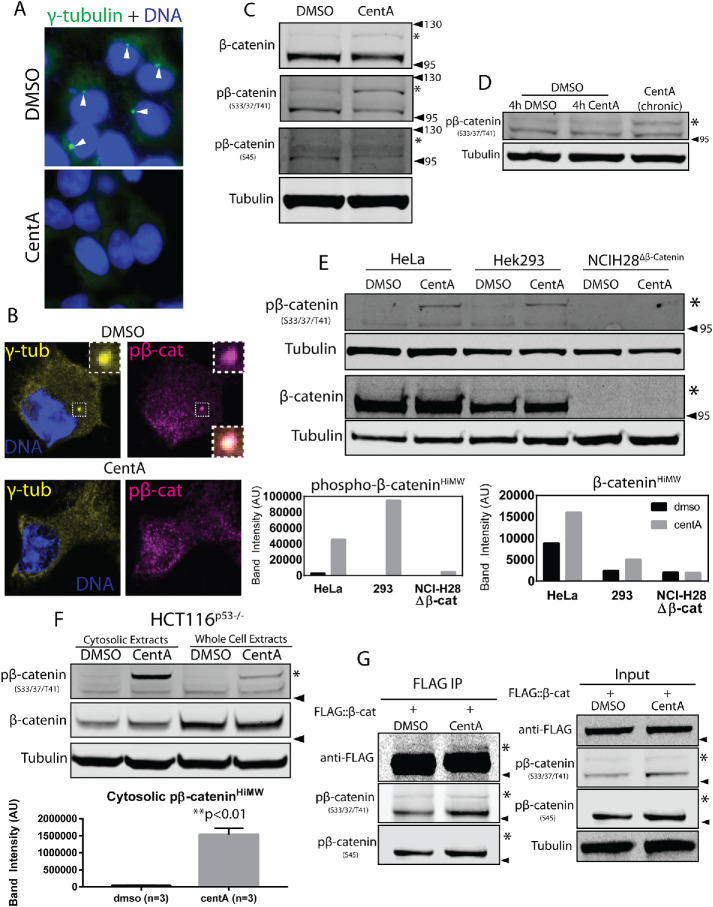
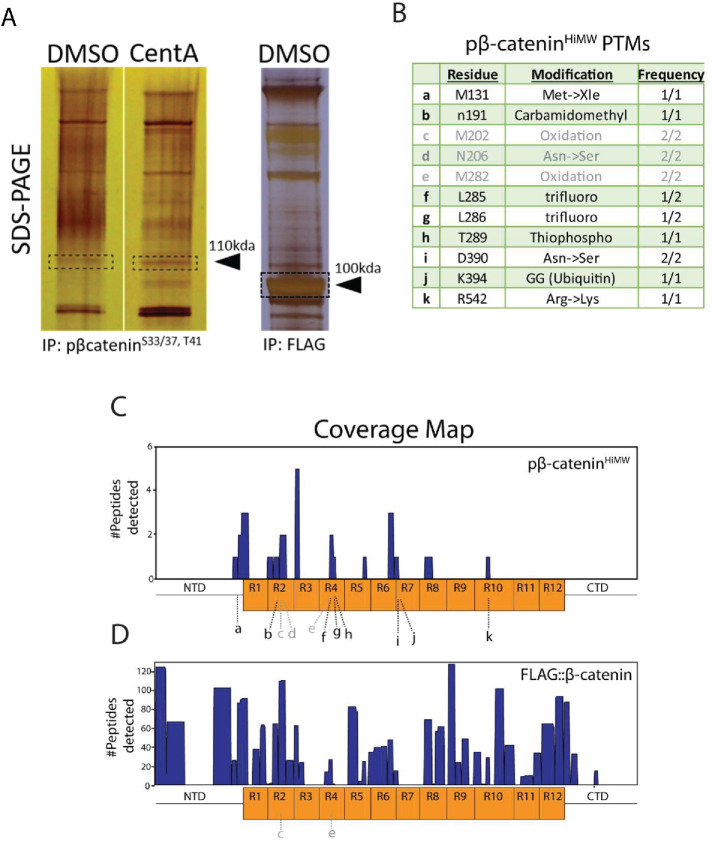
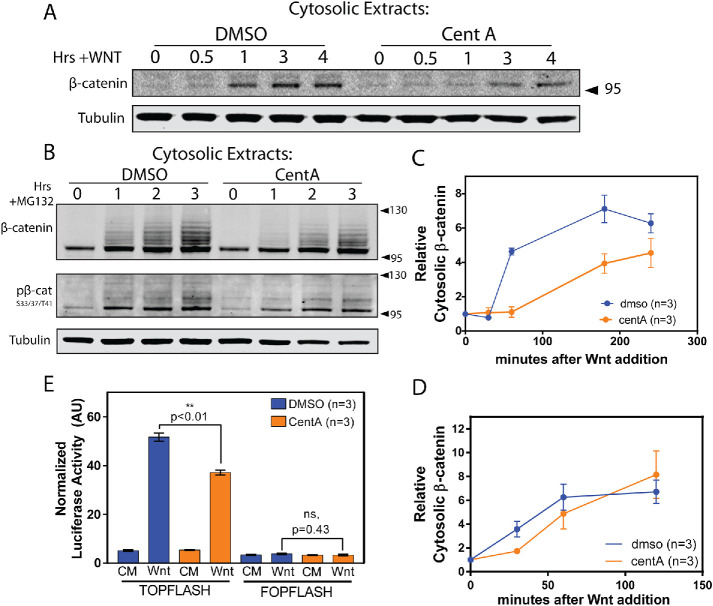
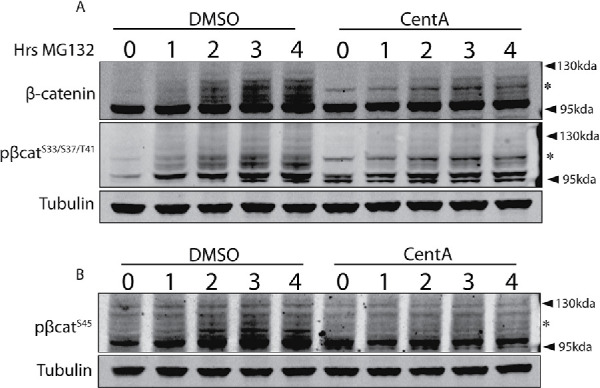
The Wnt/β-catenin signaling pathway is central to metazoan development and routinely dysregulated in cancer. Wnt/β-catenin signaling initiates transcriptional reprogramming upon stabilization of the transcription factor β-catenin, which is otherwise posttranslationally processed by a destruction complex and degraded by the proteasome. Since various Wnt signaling components are enriched at centrosomes, we examined the functional contribution of centrosomes to Wnt signaling, β-catenin regulation, and posttranslational modifications. In HEK293 cells depleted of centrosomes we find that β-catenin synthesis and degradation rates are unaffected but that the normal accumulation of β-catenin in response to Wnt signaling is attenuated. This is due to accumulation of a novel high-molecular-weight form of phosphorylated β-catenin that is constitutively degraded in the absence of Wnt. Wnt signaling operates by inhibiting the destruction complex and thereby reducing destruction complex-phosphorylated β-catenin, but high-molecular-weight β-catenin is unexpectedly increased by Wnt signaling. Therefore these studies have identified a pool of β-catenin effectively shielded from regulation by Wnt. We present a model whereby centrosomes prevent inappropriate β-catenin modifications that antagonize normal stabilization by Wnt signals.
Wnt/β-catenin 信号通路是后生动物发育的核心,并且在癌症中经常失调。Wnt/β-catenin 信号通路在转录因子 β-catenin 稳定后启动转录重编程,否则β-catenin 会被降解复合物进行翻译后加工,并被蛋白酶体降解。由于各种 Wnt 信号成分在中心体富集,我们研究了中心体对 Wnt 信号、β-catenin 调节和翻译后修饰的功能贡献。在中心体耗尽的 HEK293 细胞中,我们发现β-catenin 的合成和降解速率不受影响,但正常的 Wnt 信号诱导的β-catenin 积累受到抑制。这是由于一种新型的高相对分子质量的磷酸化β-catenin 的积累,在没有 Wnt 的情况下,它会被持续降解。Wnt 信号通过抑制破坏复合物从而减少破坏复合物磷酸化的β-catenin 来起作用,但 Wnt 信号出乎意料地增加了高分子量的β-catenin。因此,这些研究确定了一组β-catenin,它们有效地免受 Wnt 调节。我们提出了一个模型,即中心体防止了β-catenin 的不适当修饰,这些修饰拮抗了 Wnt 信号的正常稳定。