Department of Environmental Science, Policy, and Management & Museum of Vertebrate Zoology, University of California, Berkeley, CA, USA.
Department of Ecology and Evolutionary Biology & Museum of Zoology, University of Michigan, Ann Arbor, MI, USA.
BMC Evol Biol. 2020 Jul 9;20(1):80. doi: 10.1186/s12862-020-01645-0.
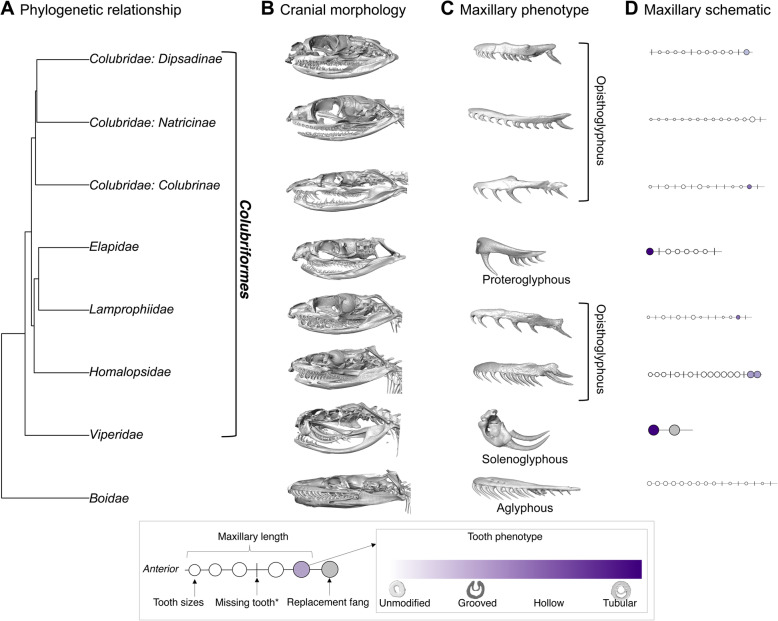
Fangs are a putative key innovation that revolutionized prey capture and feeding in snakes, and - along with their associated venom phenotypes - have made snakes perhaps the most medically-significant vertebrate animals. Three snake clades are known for their forward-positioned fangs, and these clades (Elapidae, Viperidae, and Atractaspidinae) contain the majority of snakes that are traditionally considered venomous. However, many other snakes are "rear-fanged": they possess potentially venom-delivering teeth situated at the rear end of the upper jaw. Quantification of fang phenotypes - and especially those of rear-fanged species - has proved challenging or impossible owing to the small size and relative rarity of many such snakes. Consequently, it has been difficult to understand the evolutionary history of both venom and prey-capture strategies across extant snakes. We quantified variation in the dentition of 145 colubriform ("advanced") snake species using microCT scanning and compared dental characters with ecological data on species' diet and prey capture method(s) to understand broader patterns in snake fang evolution.
Dental traits such as maxilla length, tooth number, and fang size show strong phylogenetic signal across Colubriformes. We find extreme heterogeneity and evolutionary lability in the rear-fanged phenotype in colubrid (colubrine, dipsadine, and natricine lineages) and lamprophiid snakes, in contrast to relative uniformity in the front fanged phenotypes of other groups (vipers and, to a lesser extent, elapids). Fang size and position are correlated with venom-use in vipers, elapids, and colubrid snakes, with the latter group shifting fangs anteriorly by shortening the entire maxillary bone. We find that maxilla length and tooth number may also be correlated with the evolution of dietary specialization. Finally, an ancestral state reconstruction suggests that fang loss is a recurring phenomenon in colubrid snakes, likely accompanied by shifts in diet and prey capture mode.
Our study provides a framework for quantifying the complex morphologies associated with venom use in snakes. Our results suggest that fang phenotypes, and particularly the rear-fanged phenotype, in snakes are both diverse and labile, facilitating a wide range of ecological strategies and contributing to spectacular radiations of these organisms in tropical and subtropical biomes worldwide.
毒牙是蛇类捕食和进食方式发生革命性变化的一个假定关键创新,与相关的毒液表型一起,使蛇类成为最具医学意义的脊椎动物。有三个蛇类进化支以其向前定位的毒牙而闻名,这些进化支(眼镜蛇科、蝰蛇科和响尾蛇科)包含了大多数传统上被认为是有毒的蛇类。然而,许多其他蛇类是“后齿类”:它们在上颚的后端拥有可能输送毒液的牙齿。由于许多此类蛇类的体型较小且相对罕见,因此毒牙表型的量化,特别是后齿类蛇类的毒牙表型的量化一直具有挑战性或不可能。因此,很难理解现存蛇类的毒液和捕食策略的进化历史。我们使用微 CT 扫描对 145 种游蛇类(“高级”)物种的牙齿结构进行了量化,并将牙齿特征与物种的饮食和捕食方法的生态数据进行了比较,以了解蛇类毒牙进化的更广泛模式。
牙齿特征,如上颌骨长度、牙齿数量和毒牙大小,在 Colubriformes 中具有很强的系统发育信号。我们发现,在 colubrid(colubrine、dipsadine 和 natricine 谱系)和 Lamprophiidae 蛇类中,后齿类的表型具有极强的异质性和进化不稳定性,而其他群体(蝰蛇科,在较小程度上还有眼镜蛇科)的前齿类表型则相对均匀。毒牙大小和位置与蝰蛇科、眼镜蛇科和 colubrid 蛇类的毒液使用相关,而后者通过缩短整个上颌骨使毒牙向前移动。我们发现,上颌骨长度和牙齿数量也可能与饮食特化的进化相关。最后,一个祖先状态重建表明,毒牙缺失是 colubrid 蛇类中反复出现的现象,可能伴随着饮食和捕食方式的转变。
我们的研究为量化与蛇类毒液使用相关的复杂形态提供了一个框架。我们的结果表明,蛇类的毒牙表型,特别是后齿类表型,既多样化又不稳定,这为蛇类的各种生态策略提供了便利,并促成了它们在全球热带和亚热带生物区系中的壮观辐射。