School of Medicine, University of Southampton, Southampton, UK.
Slovak Academy of Sciences, Institute of Molecular Physiology and Genetics, Bratislava, Slovak Republic.
RNA Biol. 2021 Mar;18(3):354-367. doi: 10.1080/15476286.2020.1805909. Epub 2020 Sep 23.
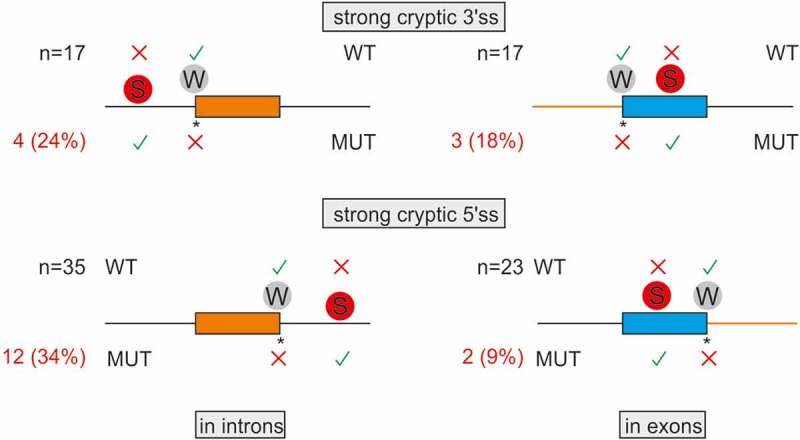
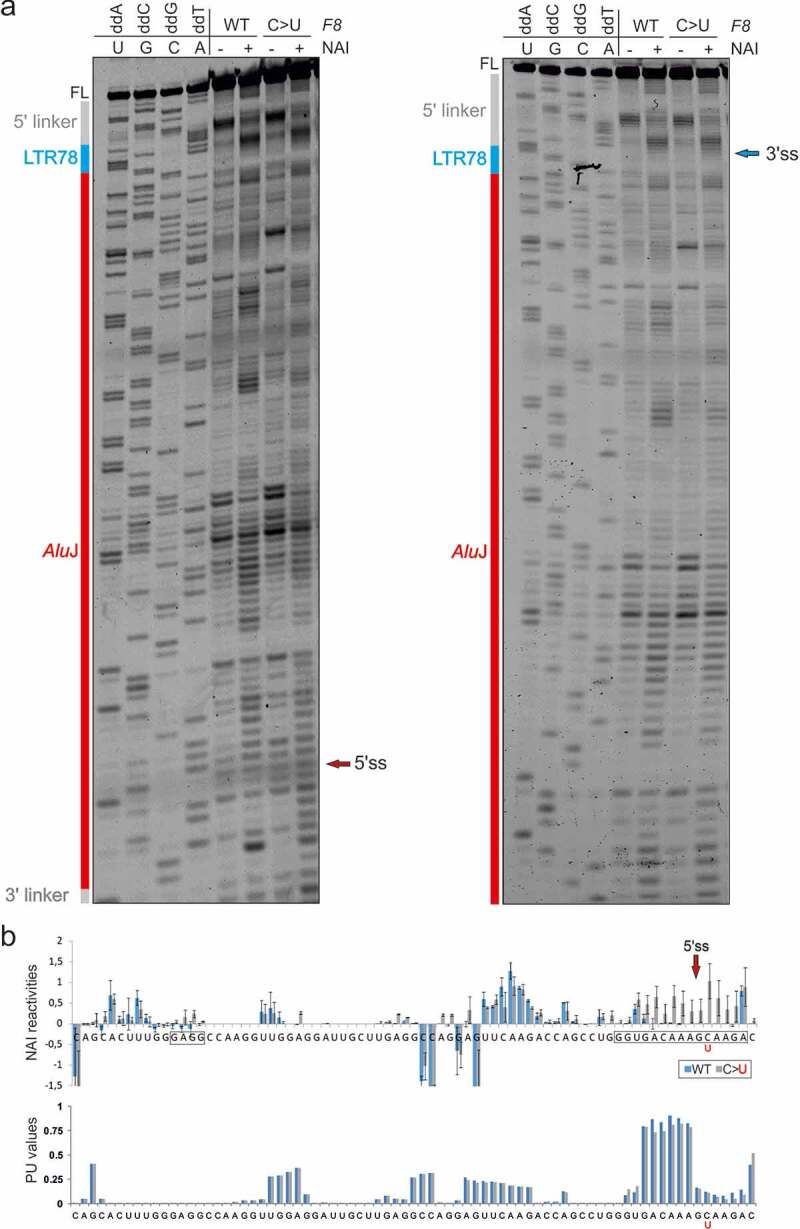
Transposed elements (TEs) have dramatically shaped evolution of the exon-intron structure and significantly contributed to morbidity, but how recent TE invasions into older TEs cooperate in generating new coding sequences is poorly understood. Employing an updated repository of new exon-intron boundaries induced by pathogenic mutations, termed DBASS, here we identify novel TE clusters that facilitated exon selection. To explore the extent to which such TE exons maintain RNA secondary structure of their progenitors, we carried out structural studies with a composite exon that was derived from a long terminal repeat (LTR78) and J and was activated by a C > T mutation optimizing the 5' splice site. Using a combination of SHAPE, DMS and enzymatic probing, we show that the disease-causing mutation disrupted a conserved J stem that evolved from helix 3.3 (or 5b) of 7SL RNA, liberating a primordial GC 5' splice site from the paired conformation for interactions with the spliceosome. The mutation also reduced flexibility of conserved residues in adjacent exon-derived loops of the central hairpin, revealing a cross-talk between traditional and auxilliary splicing motifs that evolved from opposite termini of 7SL RNA and were approximated by Watson-Crick base-pairing already in organisms without spliceosomal introns. We also identify existing exons activated by the same RNA rearrangement. Collectively, these results provide valuable TE exon models for studying formation and kinetics of pre-mRNA building blocks required for splice-site selection and will be useful for fine-tuning auxilliary splicing motifs and exon and intron size constraints that govern aberrant splice-site activation.
转座元件 (TEs) 极大地影响了外显子-内含子结构的进化,并显著导致了发病率,但最近 TE 对旧 TE 的入侵如何协同产生新的编码序列仍知之甚少。利用一个更新的致病性突变诱导的新外显子-内含子边界的存储库,称为 DBASS,我们在这里确定了有助于外显子选择的新 TE 簇。为了探索这些 TE 外显子在多大程度上保持其前体的 RNA 二级结构,我们对一个源自长末端重复序列 (LTR78) 和 J 的复合外显子进行了结构研究,该外显子由一个 C>T 突变激活,优化了 5' 剪接位点。我们使用 SHAPE、DMS 和酶探测的组合,表明致病突变破坏了一个保守的 J 茎,该茎是从 7SL RNA 的 3.3 螺旋(或 5b)进化而来的,将原始 GC 5' 剪接位点从配对构象中释放出来,与剪接体相互作用。该突变还降低了中央发夹中相邻外显子衍生环中保守残基的灵活性,揭示了来自 7SL RNA 相反末端的传统和辅助剪接基序之间的交叉对话,这些基序已经在没有剪接体内含子的生物体中通过 Watson-Crick 碱基配对而接近。我们还鉴定了由相同 RNA 重排激活的现有外显子。总的来说,这些结果为研究剪接位点选择所需的 pre-mRNA 构建块的形成和动力学提供了有价值的 TE 外显子模型,并将有助于微调辅助剪接基序以及外显子和内含子大小约束,这些约束控制着异常剪接位点的激活。