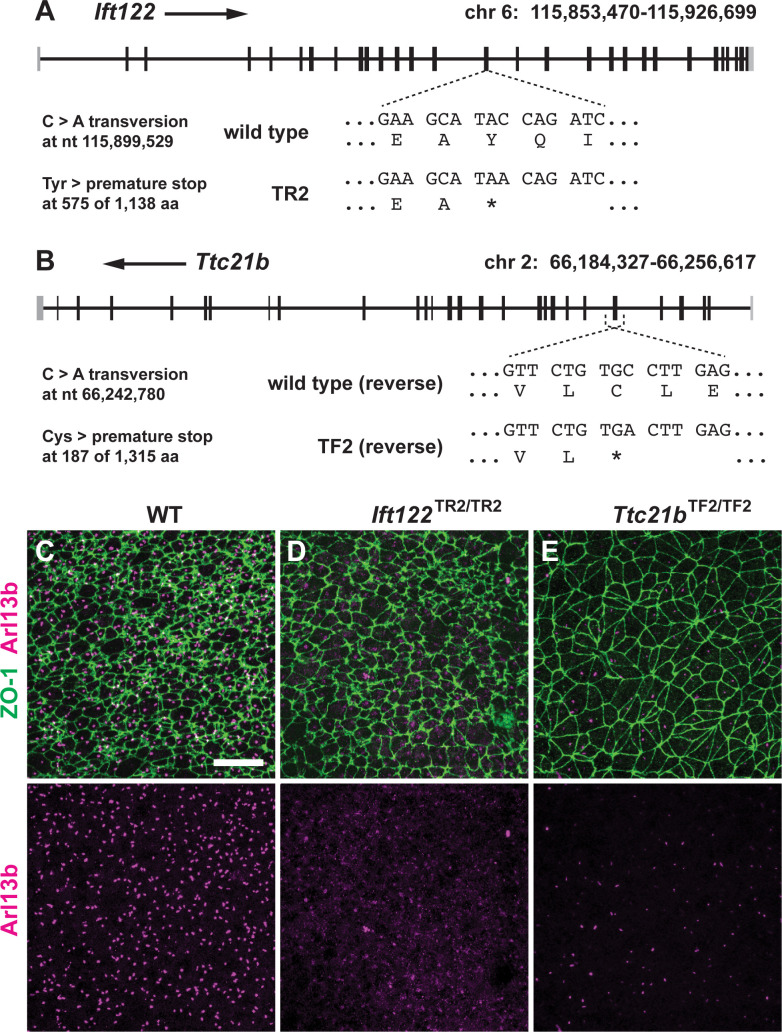
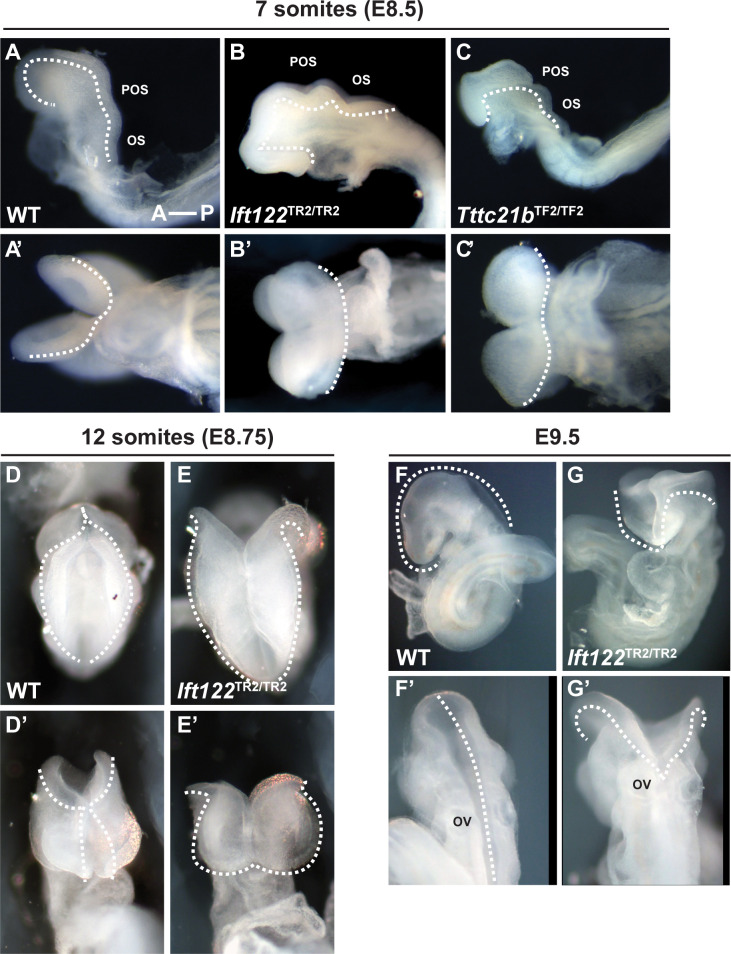
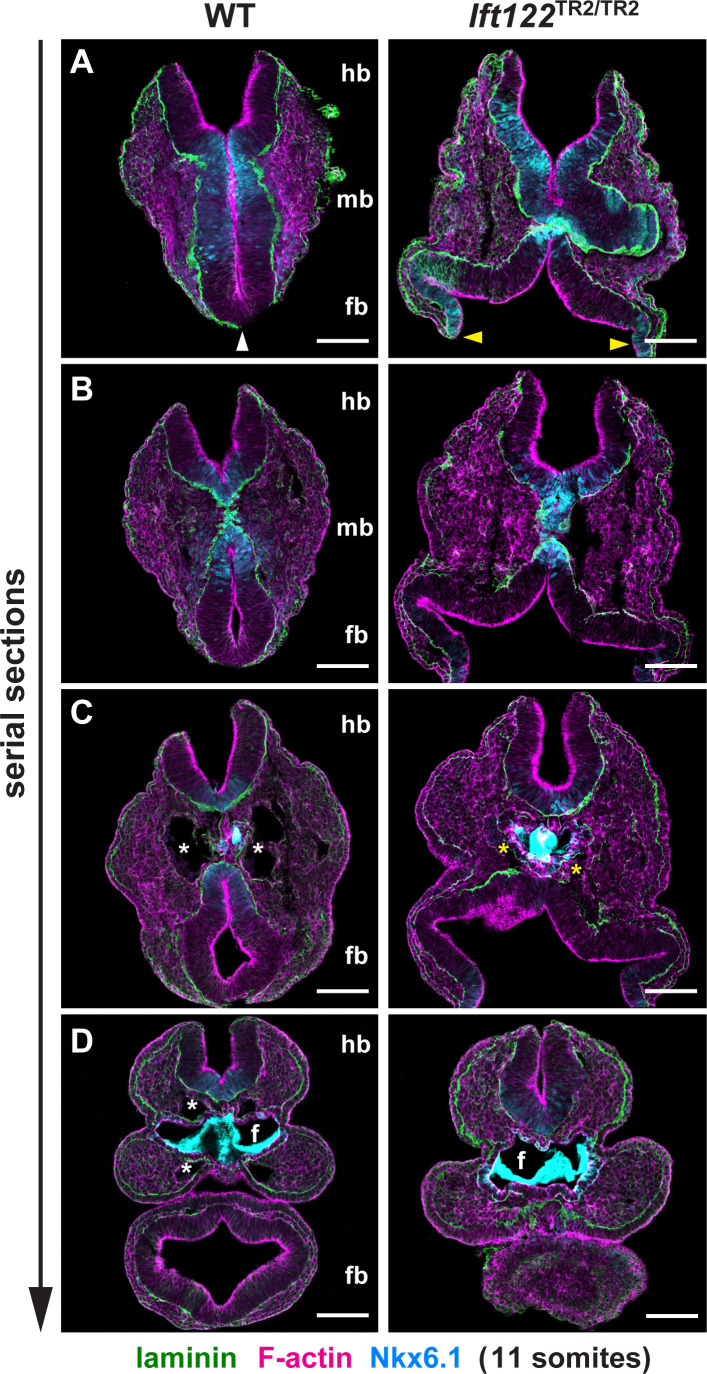
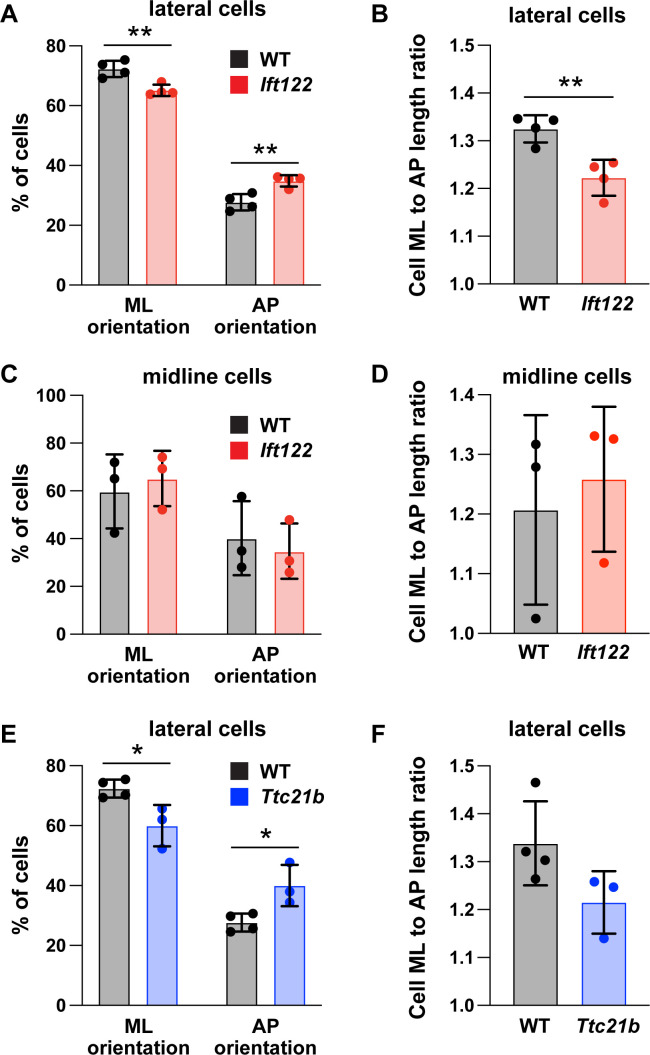
Howard Hughes Medical Institute and Developmental Biology Program, Sloan Kettering Institute, New York, United States.
Developmental Biology Program, Sloan Kettering Institute, New York, United States.
Elife. 2020 Oct 26;9:e60234. doi: 10.7554/eLife.60234.
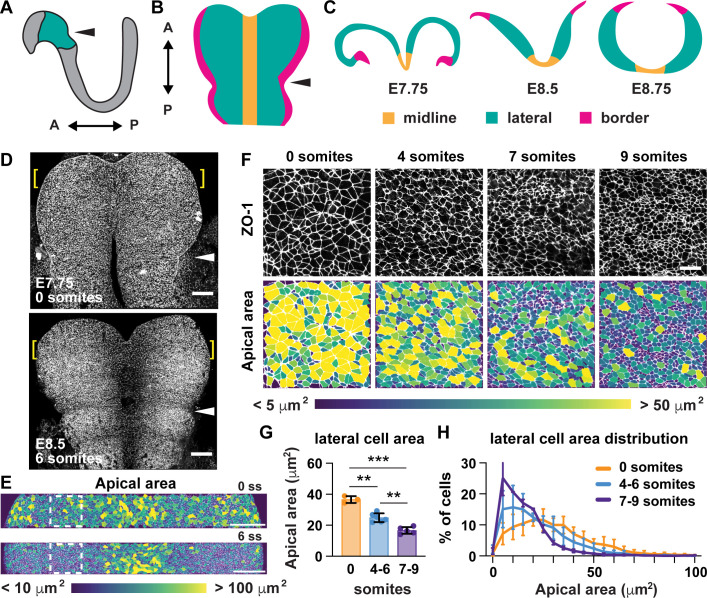
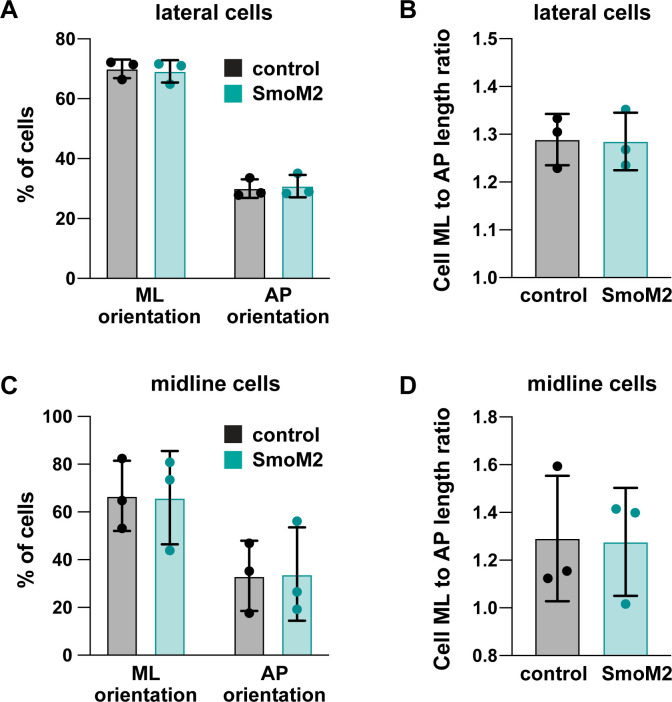
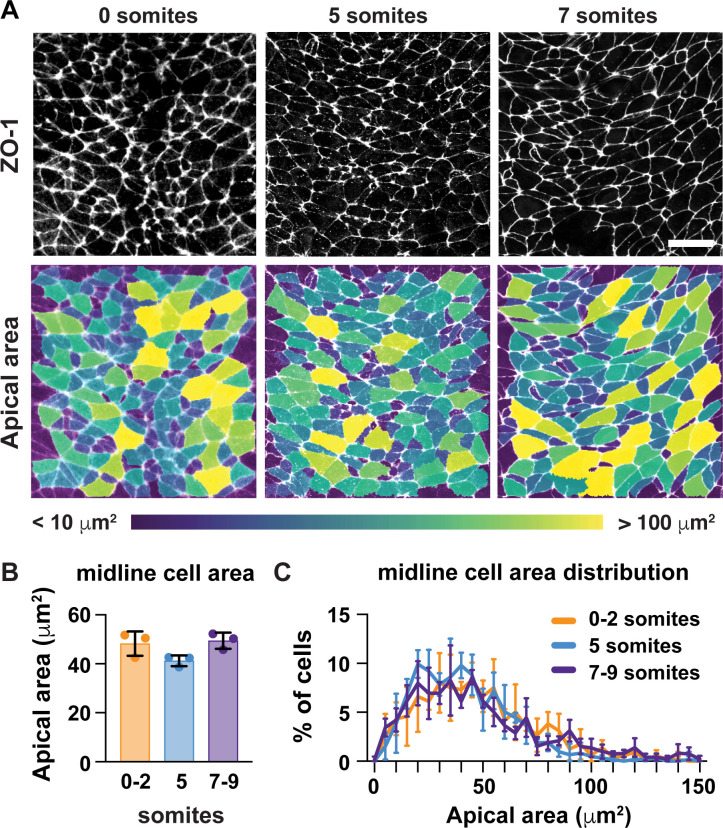
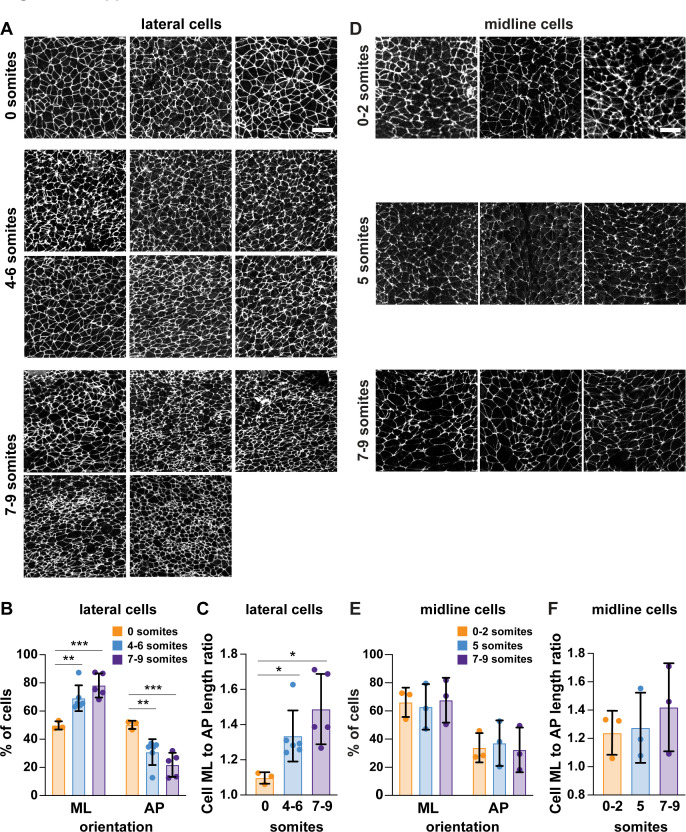
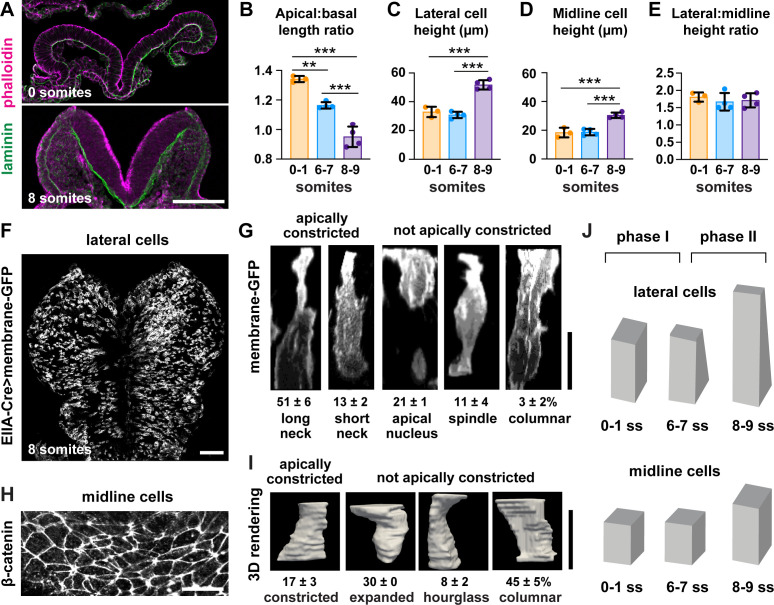
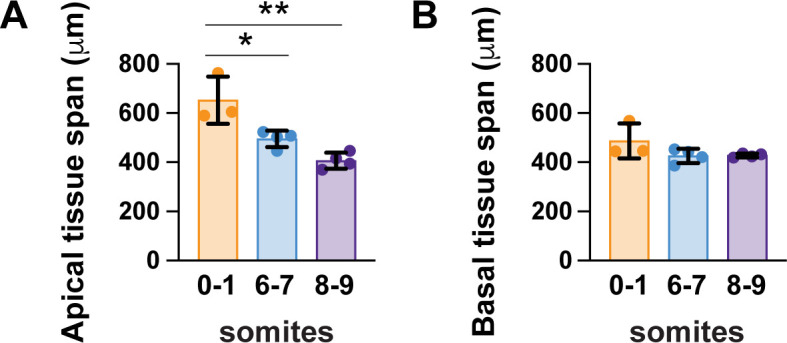
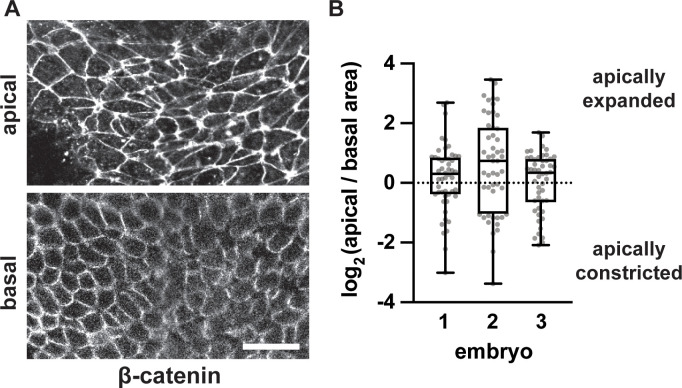
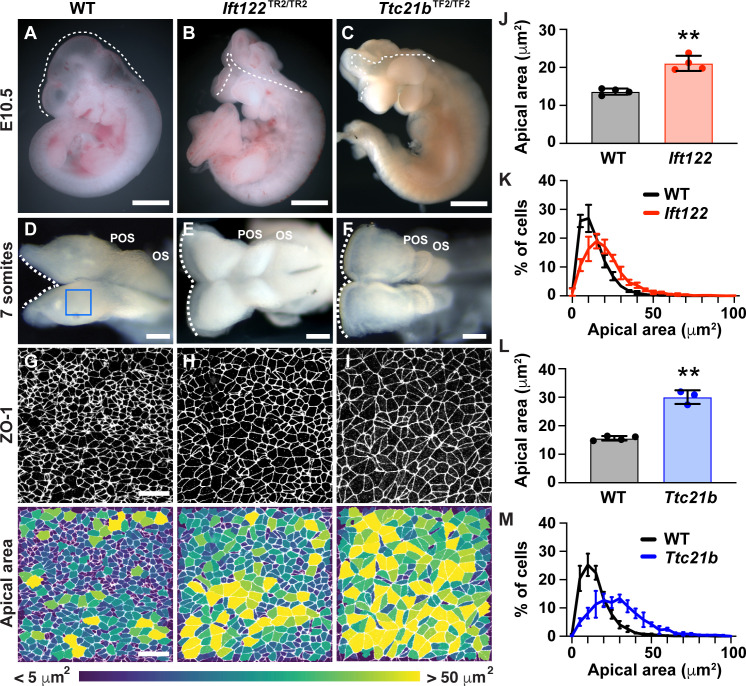
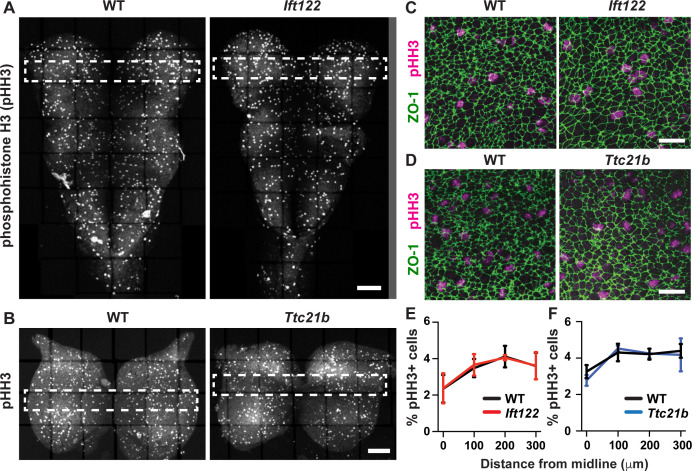
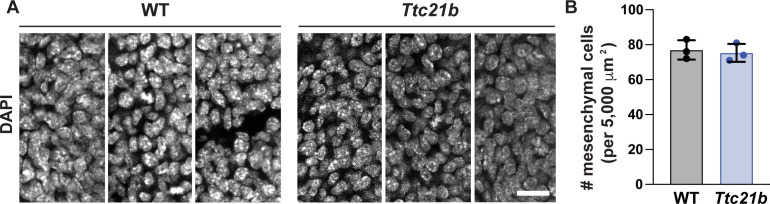
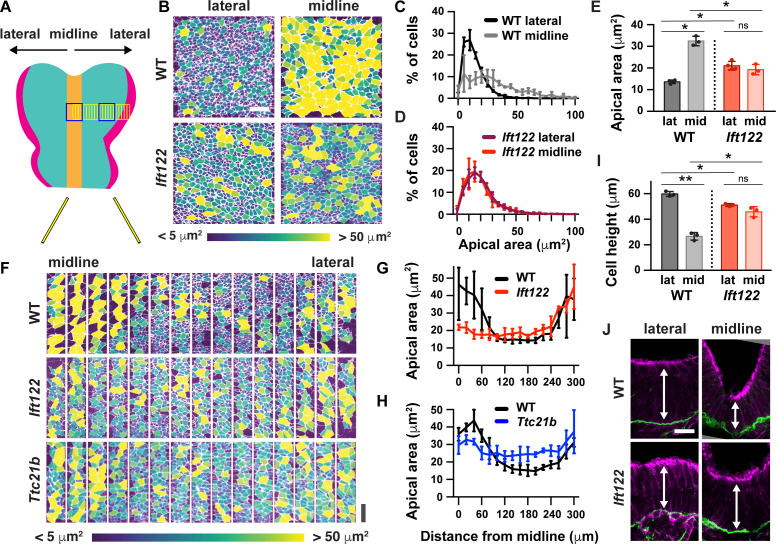
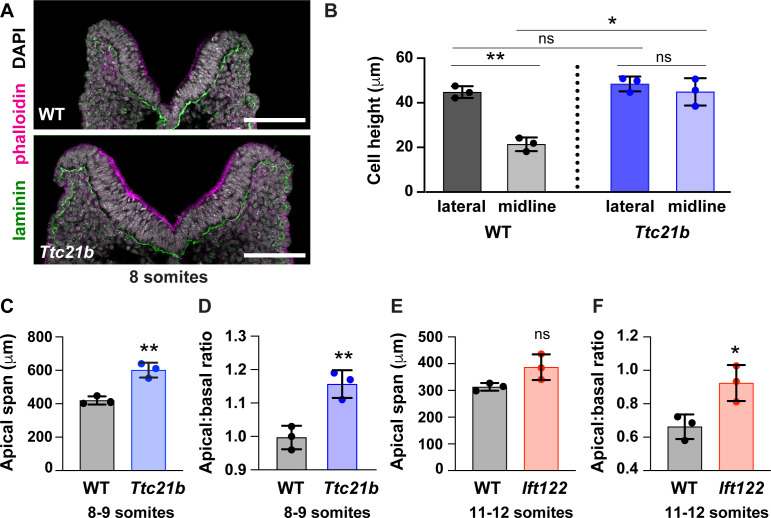
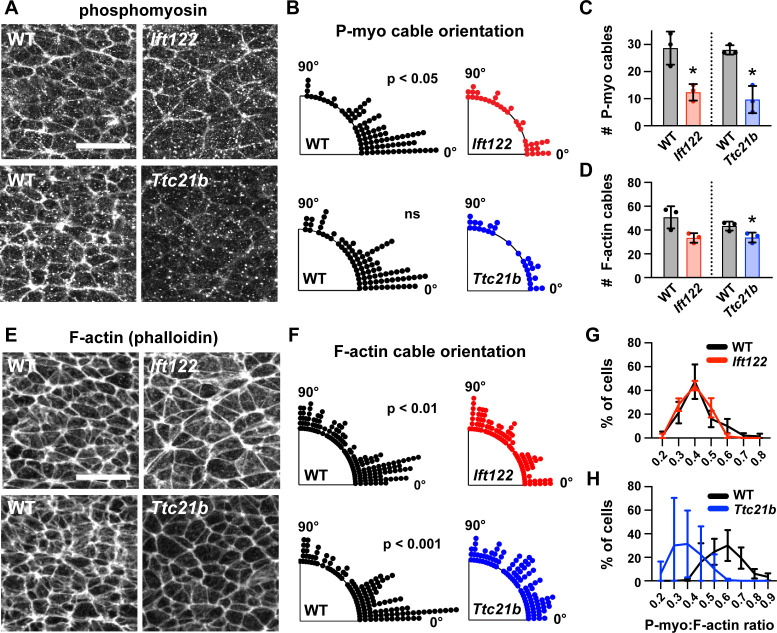
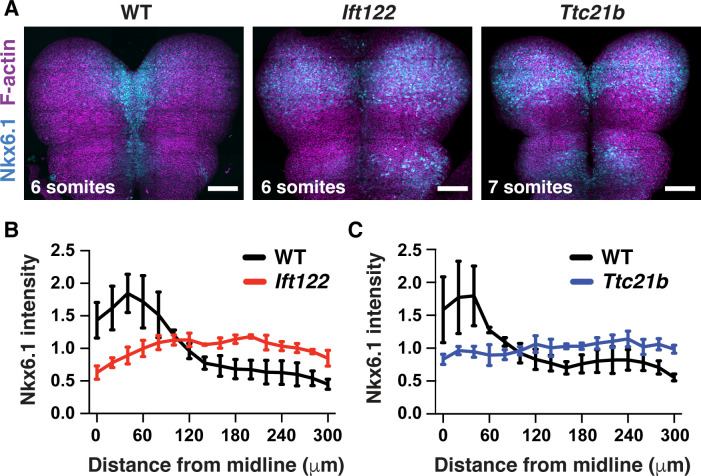
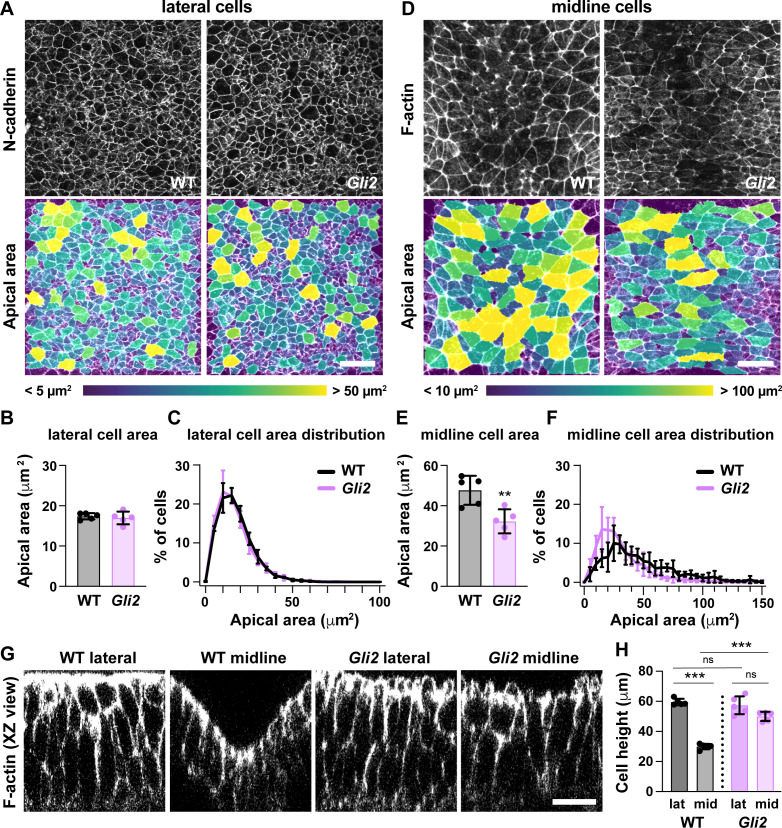
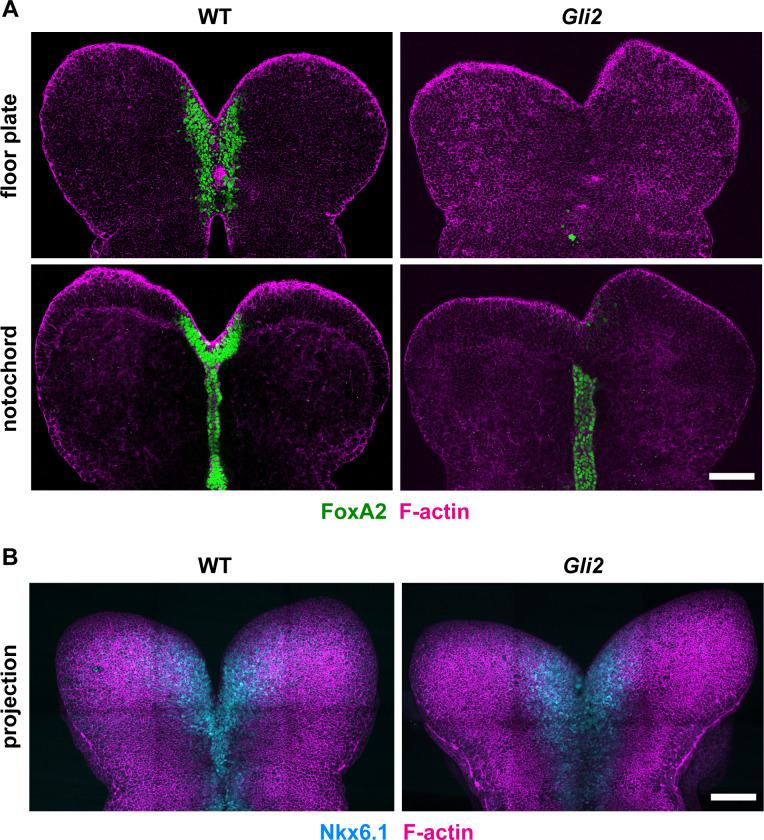
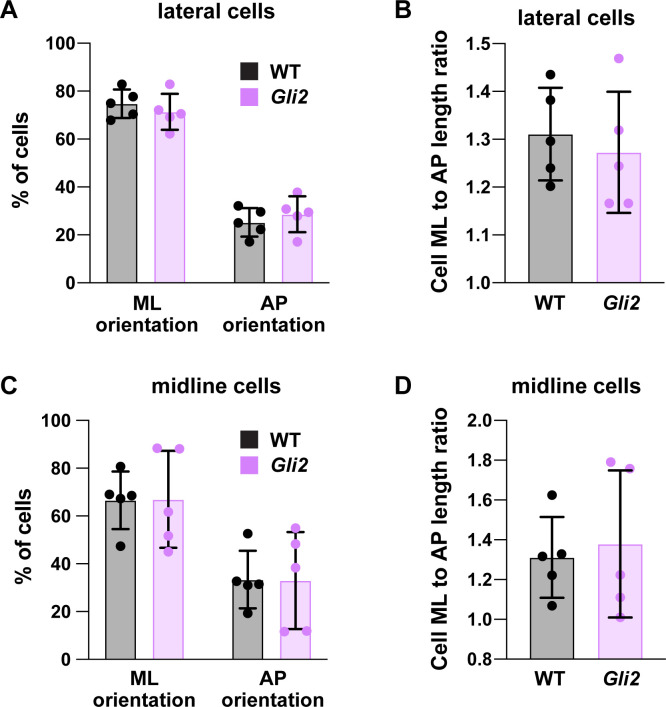
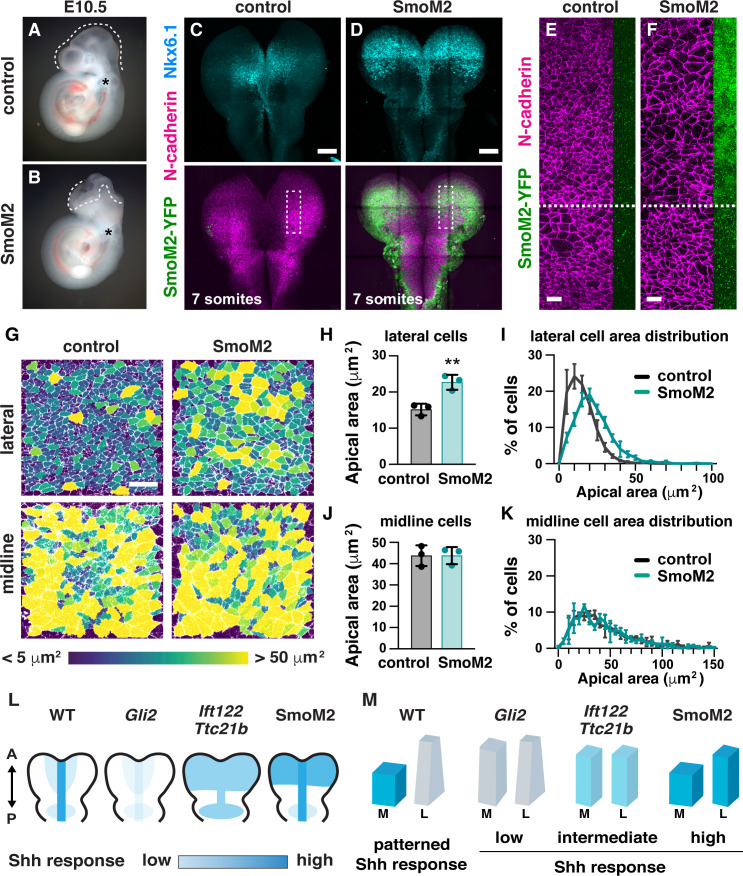
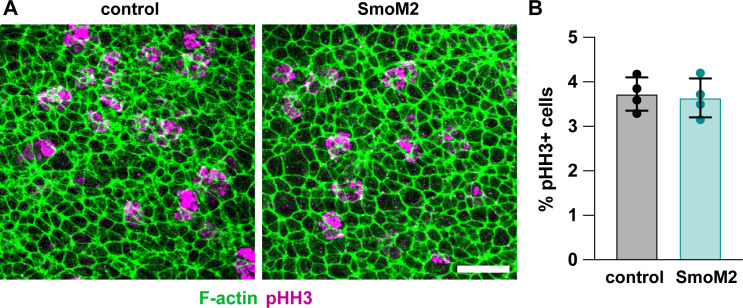
Neural tube closure defects are a major cause of infant mortality, with exencephaly accounting for nearly one-third of cases. However, the mechanisms of cranial neural tube closure are not well understood. Here, we show that this process involves a tissue-wide pattern of apical constriction controlled by Sonic hedgehog (Shh) signaling. Midline cells in the mouse midbrain neuroepithelium are flat with large apical surfaces, whereas lateral cells are taller and undergo synchronous apical constriction, driving neural fold elevation. Embryos lacking the Shh effector Gli2 fail to produce appropriate midline cell architecture, whereas embryos with expanded Shh signaling, including the IFT-A complex mutants and and embryos expressing activated Smoothened, display apical constriction defects in lateral cells. Disruption of lateral, but not midline, cell remodeling results in exencephaly. These results reveal a morphogenetic program of patterned apical constriction governed by Shh signaling that generates structural changes in the developing mammalian brain.
神经管闭合缺陷是婴儿死亡的主要原因,其中无脑畸形约占三分之一。然而,颅神经管闭合的机制尚不清楚。在这里,我们表明这个过程涉及 Sonic hedgehog(Shh)信号控制的全组织顶壁收缩模式。小鼠中脑神经上皮的中线细胞是扁平的,具有较大的顶表面,而侧部细胞较高,并经历同步的顶壁收缩,从而驱动神经褶升高。缺乏 Shh 效应物 Gli2 的胚胎无法产生适当的中线细胞结构,而具有扩展的 Shh 信号的胚胎,包括 IFT-A 复合物突变体 和 以及表达激活型 Smoothened 的胚胎,在侧部细胞中表现出顶壁收缩缺陷。侧部但不是中线细胞重塑的破坏导致无脑畸形。这些结果揭示了由 Shh 信号控制的模式化顶壁收缩的形态发生程序,该程序在发育中的哺乳动物大脑中产生结构变化。