Kyoto University, Graduate School of Biostudies, Japan.
Department of Chemistry and Biochemistry, University of California, San Diego, United States of America.
PLoS Genet. 2020 Nov 11;16(11):e1008968. doi: 10.1371/journal.pgen.1008968. eCollection 2020 Nov.
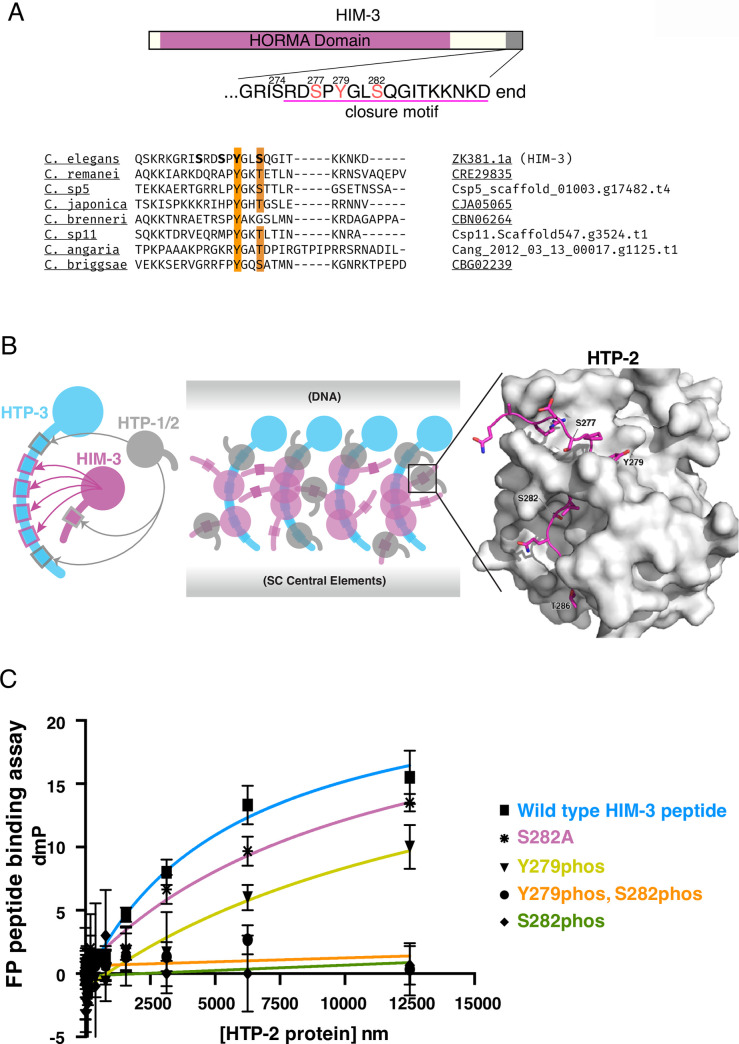
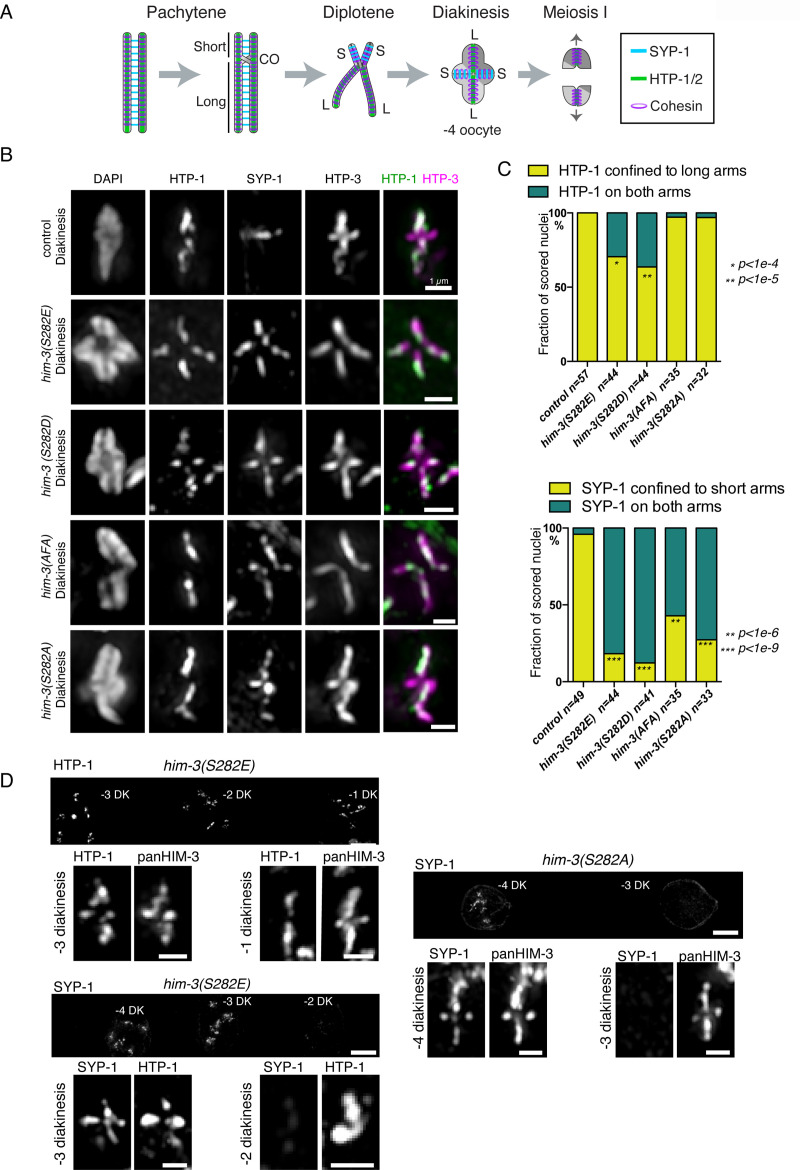
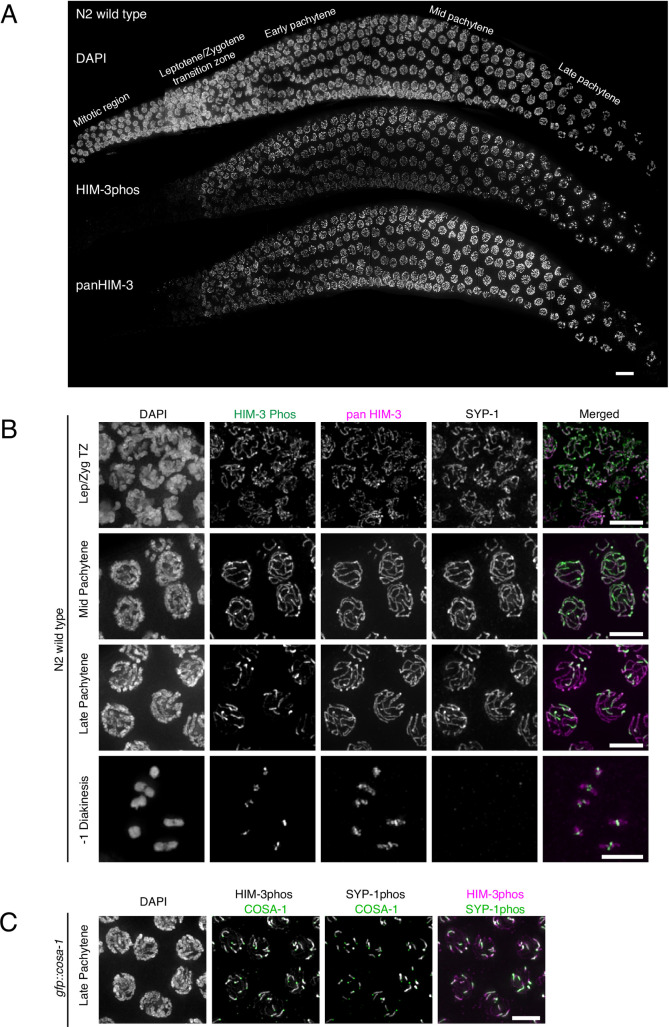
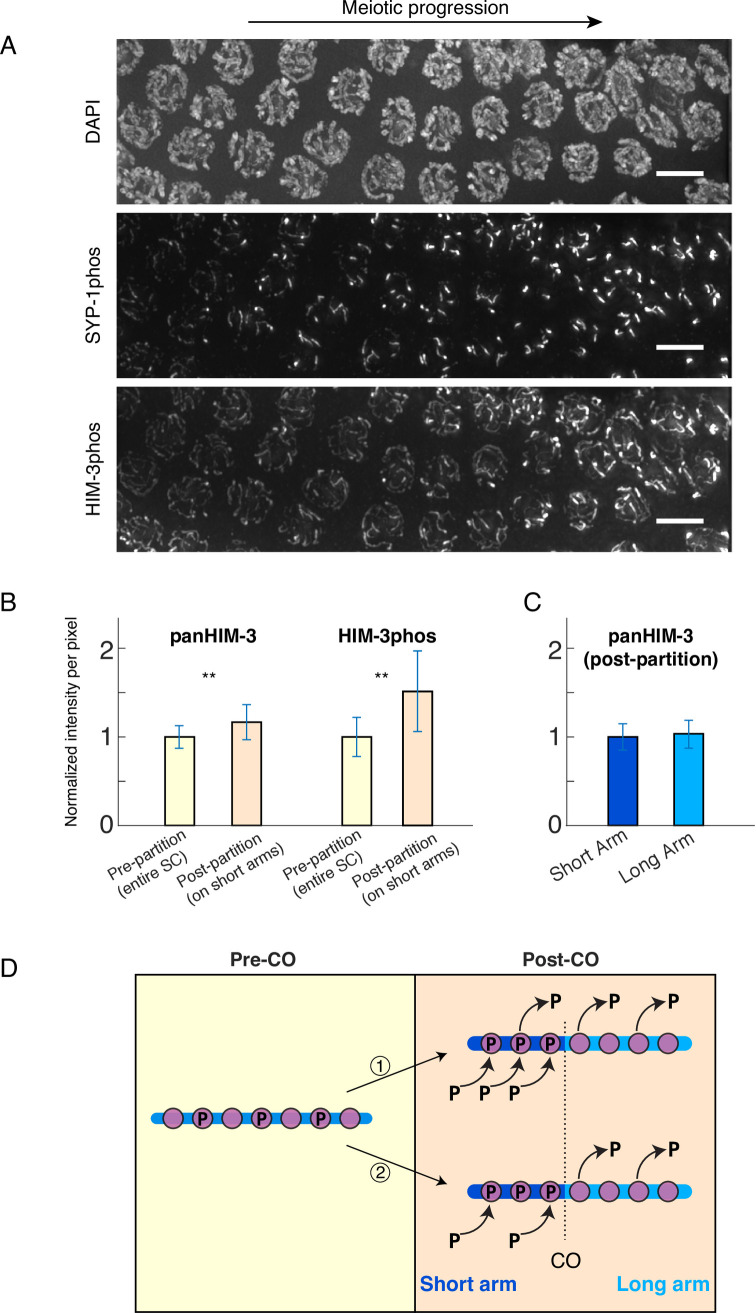
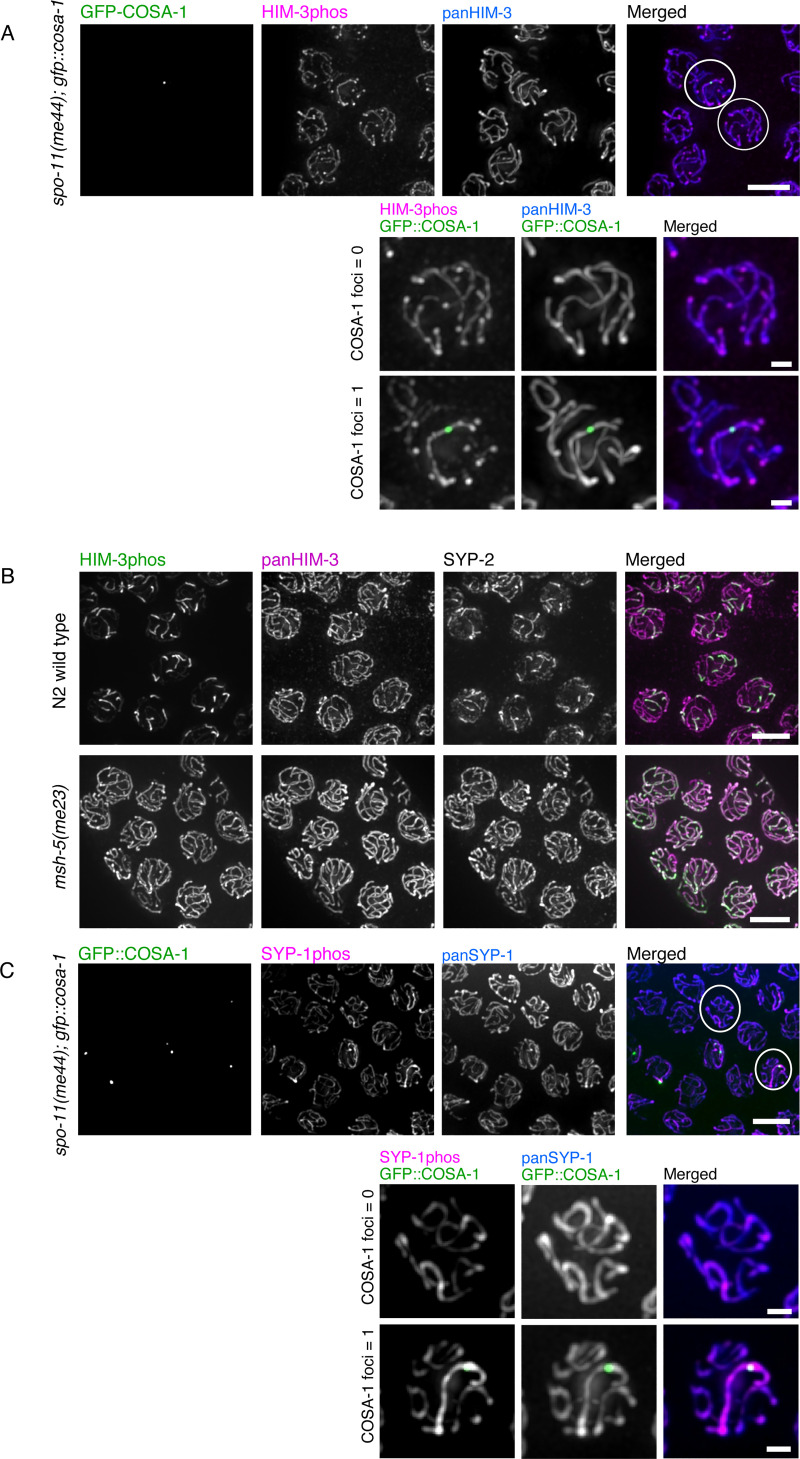
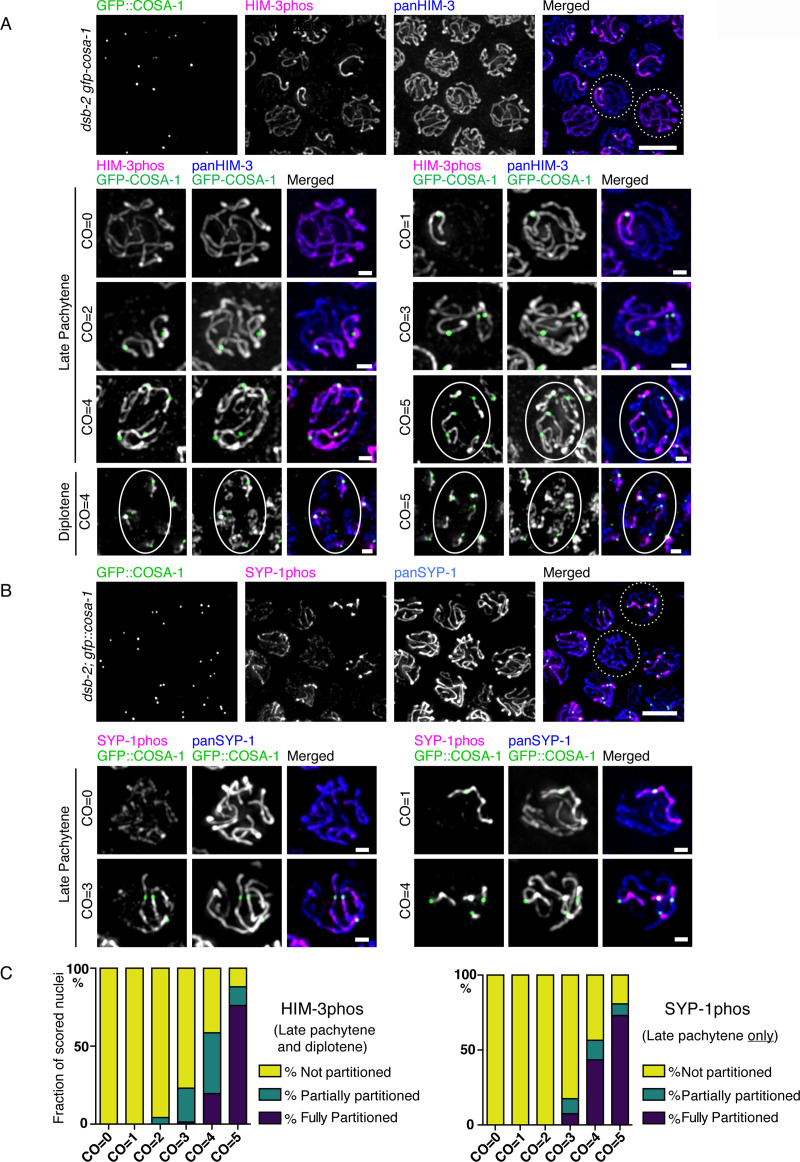
In the two cell divisions of meiosis, diploid genomes are reduced into complementary haploid sets through the discrete, two-step removal of chromosome cohesion, a task carried out in most eukaryotes by protecting cohesion at the centromere until the second division. In eukaryotes without defined centromeres, however, alternative strategies have been innovated. The best-understood of these is found in the nematode Caenorhabditis elegans: after the single off-center crossover divides the chromosome into two segments, or arms, several chromosome-associated proteins or post-translational modifications become specifically partitioned to either the shorter or longer arm, where they promote the correct timing of cohesion loss through as-yet unknown mechanisms. Here, we investigate the meiotic axis HORMA-domain protein HIM-3 and show that it becomes phosphorylated at its C-terminus, within the conserved "closure motif" region bound by the related HORMA-domain proteins HTP-1 and HTP-2. Binding of HTP-2 is abrogated by phosphorylation of the closure motif in in vitro assays, strongly suggesting that in vivo phosphorylation of HIM-3 likely modulates the hierarchical structure of the chromosome axis. Phosphorylation of HIM-3 only occurs on synapsed chromosomes, and similarly to other previously-described phosphorylated proteins of the synaptonemal complex, becomes restricted to the short arm after designation of crossover sites. Regulation of HIM-3 phosphorylation status is required for timely disassembly of synaptonemal complex central elements from the long arm, and is also required for proper timing of HTP-1 and HTP-2 dissociation from the short arm. Phosphorylation of HIM-3 thus plays a role in establishing the identity of short and long arms, thereby contributing to the robustness of the two-step chromosome segregation.
在减数分裂的两个细胞分裂过程中,通过染色体凝聚的离散两步去除,将二倍体基因组减少为互补的单倍体组,这一任务在大多数真核生物中通过保护着丝粒处的凝聚来完成,直到第二次分裂。然而,在没有定义着丝粒的真核生物中,已经创新了替代策略。其中最被理解的是在线虫 Caenorhabditis elegans 中发现的:在单个偏离中心的交叉将染色体分为两个片段或臂之后,几种染色体相关蛋白或翻译后修饰特异性分配到较短或较长的臂,在那里它们通过尚未知的机制促进凝聚丢失的正确时机。在这里,我们研究了减数分裂轴 HORMA 结构域蛋白 HIM-3,并表明它在其 C 末端发生磷酸化,在保守的“封闭基序”区域内,该区域由相关的 HORMA 结构域蛋白 HTP-1 和 HTP-2 结合。在体外测定中,封闭基序的磷酸化会破坏 HTP-2 的结合,强烈表明 HIM-3 的体内磷酸化可能调节染色体轴的层次结构。HIM-3 的磷酸化仅发生在联会染色体上,并且与其他先前描述的联会复合体磷酸化蛋白类似,在交叉位点指定后,它仅限制在短臂上。HIM-3 磷酸化状态的调节对于从长臂及时拆卸联会复合体中心元件是必需的,并且对于 HTP-1 和 HTP-2 从短臂的正确释放也是必需的。因此,HIM-3 的磷酸化在建立短臂和长臂的身份方面发挥作用,从而有助于两步染色体分离的稳健性。