Division of Biology and Biological Engineering, California Institute of Technology, Pasadena, California, USA.
Division of Biology and Biological Engineering, California Institute of Technology, Pasadena, California, USA
J Bacteriol. 2021 Jan 11;203(3). doi: 10.1128/JB.00576-20.
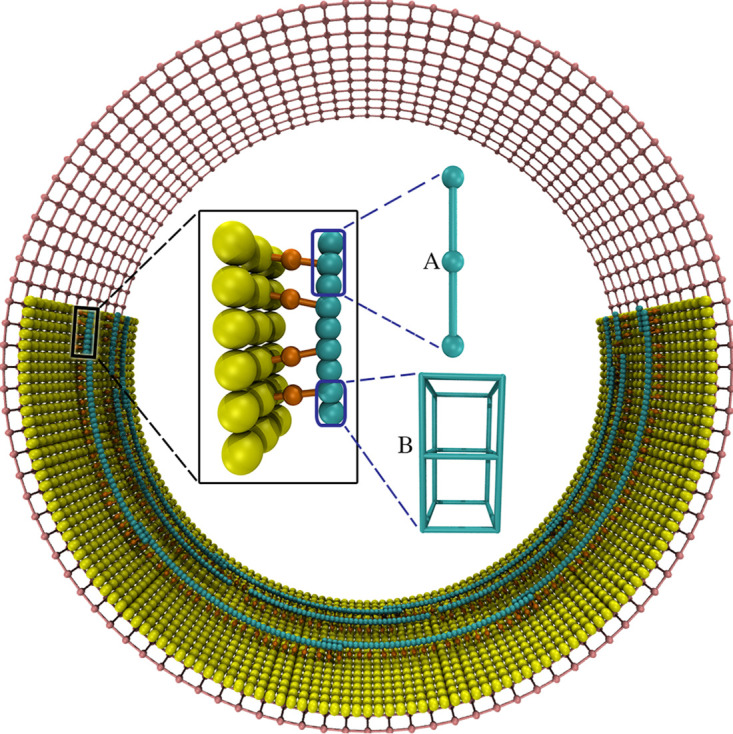
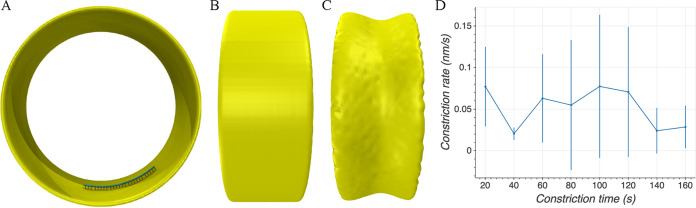
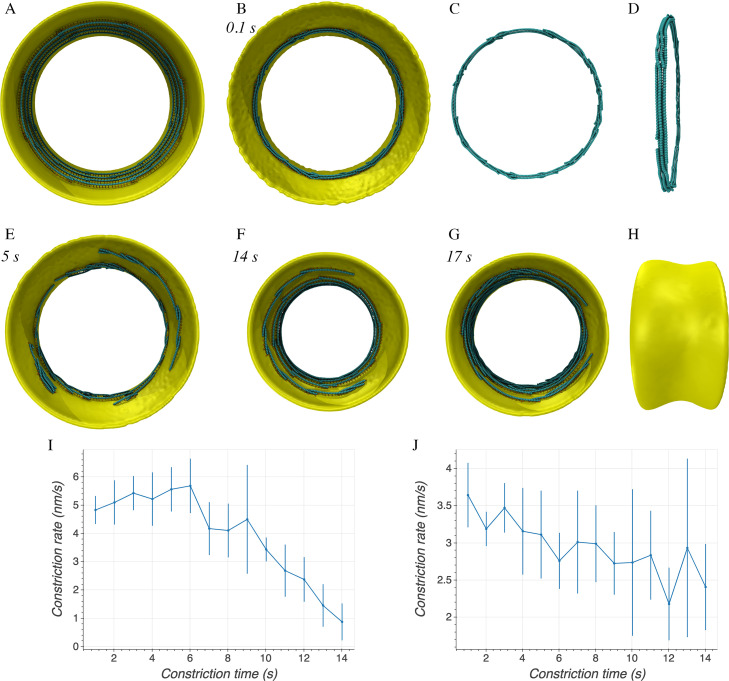
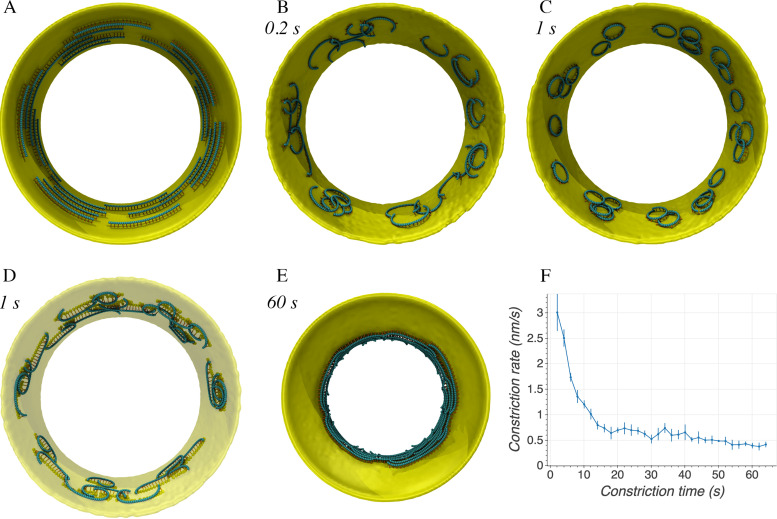
To divide, bacteria must constrict their membranes against significant force from turgor pressure. A tubulin homolog, FtsZ, is thought to drive constriction, but how FtsZ filaments might generate constrictive force in the absence of motor proteins is not well understood. There are two predominant models in the field. In one, FtsZ filaments overlap to form complete rings around the circumference of the cell, and attractive forces cause filaments to slide past each other to maximize lateral contact. In the other, filaments exert force on the membrane by a GTP-hydrolysis-induced switch in conformation from straight to bent. Here, we developed software, ZCONSTRICT, for quantitative three-dimensional (3D) simulations of Gram-negative bacterial cell division to test these two models and identify critical conditions required for them to work. We find that the avidity of any kind of lateral interactions quickly halts the sliding of filaments, so a mechanism such as depolymerization or treadmilling is required to sustain constriction by filament sliding. For filament bending, we find that a mechanism such as the presence of a rigid linker is required to constrain bending to within the division plane and maintain the distance observed between the filaments and the membrane. Of these two models, only the filament bending model is consistent with our lab's recent observation of constriction associated with a single, short FtsZ filament. FtsZ is thought to generate constrictive force to divide the cell, possibly via one of two predominant models in the field. In one, FtsZ filaments overlap to form complete rings which constrict as filaments slide past each other to maximize lateral contact. In the other, filaments exert force on the membrane by switching conformation from straight to bent. Here, we developed software, ZCONSTRICT, for three-dimensional (3D) simulations to test these two models. We find that a mechanism such as depolymerization or treadmilling are required to sustain constriction by filament sliding. For filament bending, we find that a mechanism that constrains bending to within the division plane is required to maintain the distance observed between the filaments and the membrane.
为了进行分裂,细菌必须在渗透压的巨大压力下收缩其膜。微管同源物 FtsZ 被认为可以驱动收缩,但在没有马达蛋白的情况下,FtsZ 丝如何产生收缩力还不是很清楚。该领域有两种主要的模型。在一种模型中,FtsZ 丝重叠形成完整的环围绕细胞的周长,吸引力导致丝相互滑动以最大限度地增加侧向接触。在另一种模型中,丝通过构象从直变弯的 GTP 水解诱导开关对膜施加力。在这里,我们开发了软件 ZCONSTRICT,用于革兰氏阴性细菌细胞分裂的定量三维 (3D) 模拟,以测试这两种模型,并确定它们发挥作用所需的关键条件。我们发现任何类型的侧向相互作用的亲合力都会迅速停止丝的滑动,因此需要解聚或履带式运动等机制来维持丝滑动的收缩。对于丝弯曲,我们发现需要刚性连接体的存在等机制来将弯曲限制在分裂平面内并保持观察到的丝与膜之间的距离。在这两种模型中,只有丝弯曲模型与我们实验室最近观察到的与单个短 FtsZ 丝相关的收缩一致。FtsZ 被认为通过两种主要模型之一产生收缩力来分裂细胞。在一种模型中,FtsZ 丝重叠形成完整的环,当丝相互滑动以最大限度地增加侧向接触时,环会收缩。在另一种模型中,丝通过从直变弯的构象开关对膜施加力。在这里,我们开发了软件 ZCONSTRICT,用于三维 (3D) 模拟来测试这两种模型。我们发现需要解聚或履带式运动等机制来维持丝滑动的收缩。对于丝弯曲,我们发现需要将弯曲限制在分裂平面内的机制来保持观察到的丝与膜之间的距离。