Department of Immunology, The University of Texas MD Anderson Cancer Center, Houston, TX, USA.
National Creative Research Initiatives Center for Adipose Tissue Remodeling, Department of Biological Sciences, Institute of Molecular Biology and Genetics, Seoul National University, Seoul, Korea.
Nat Immunol. 2021 Feb;22(2):193-204. doi: 10.1038/s41590-020-00829-6. Epub 2021 Jan 4.
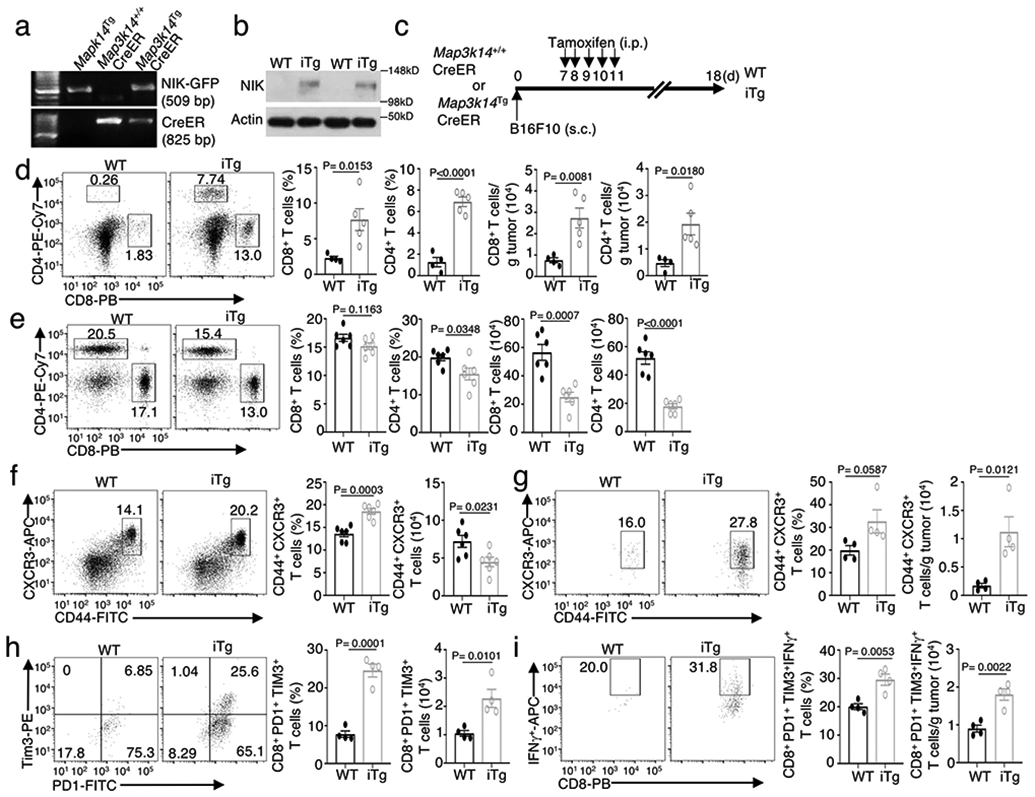
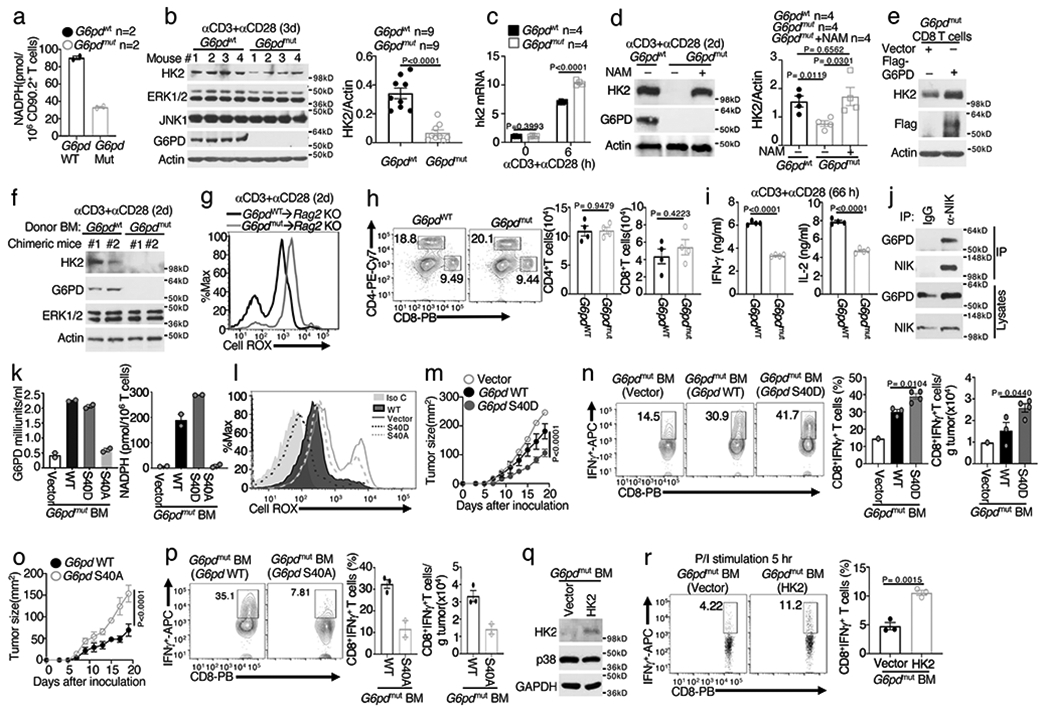
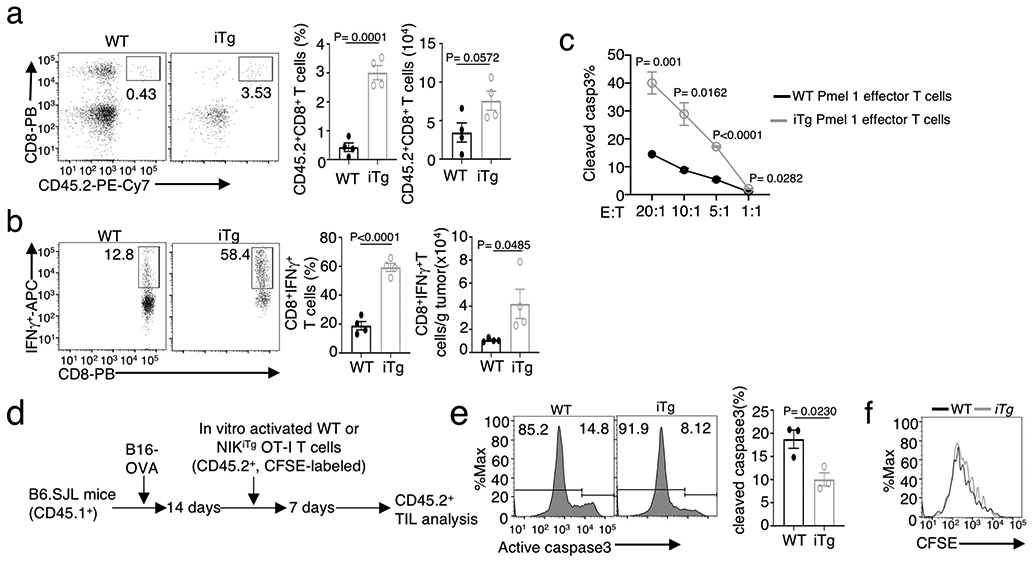
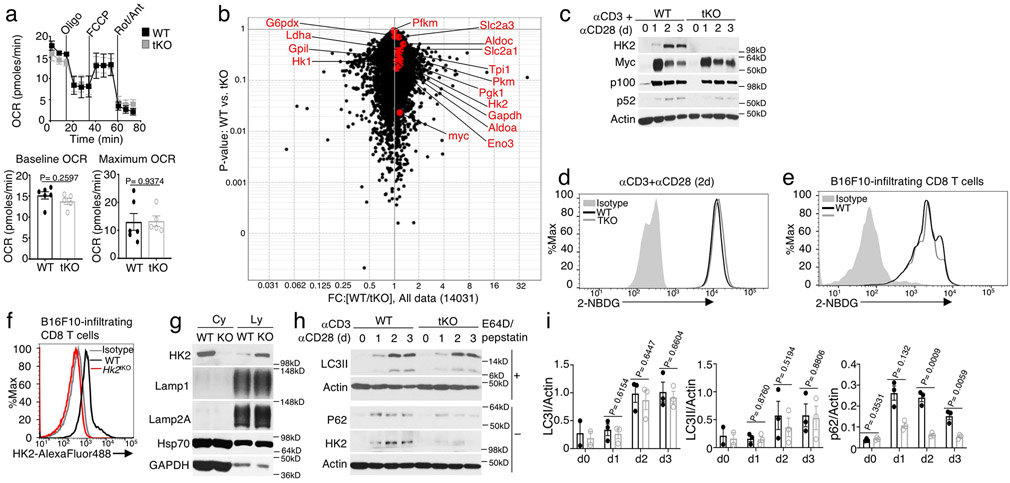
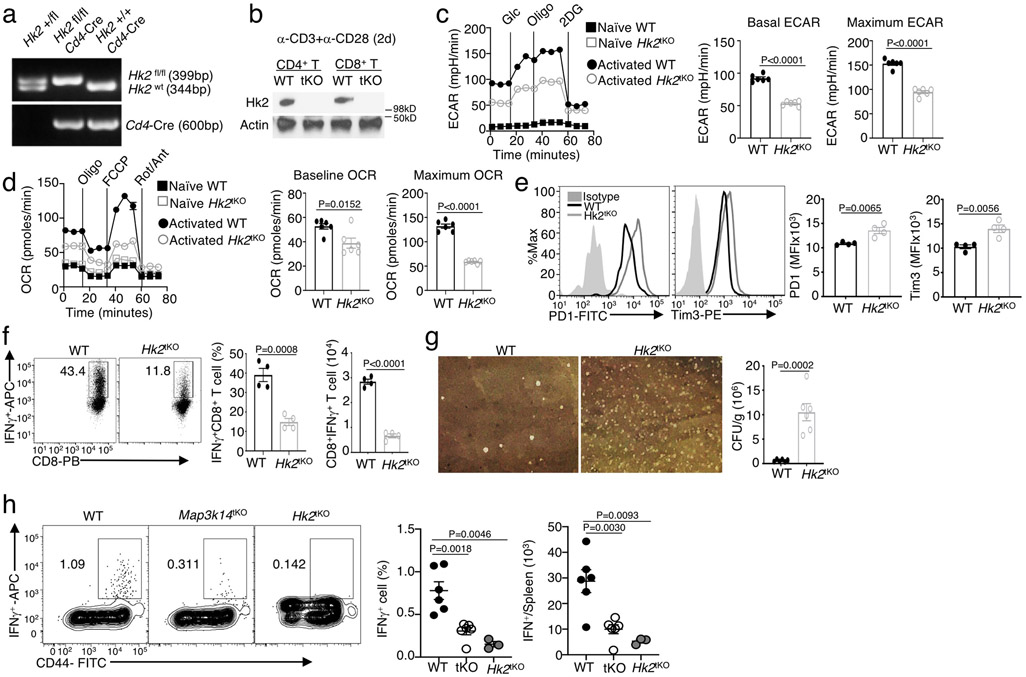
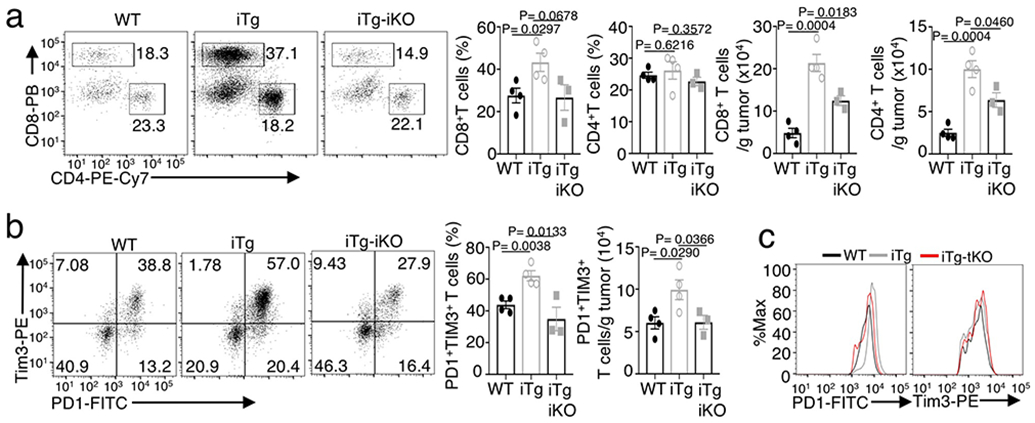
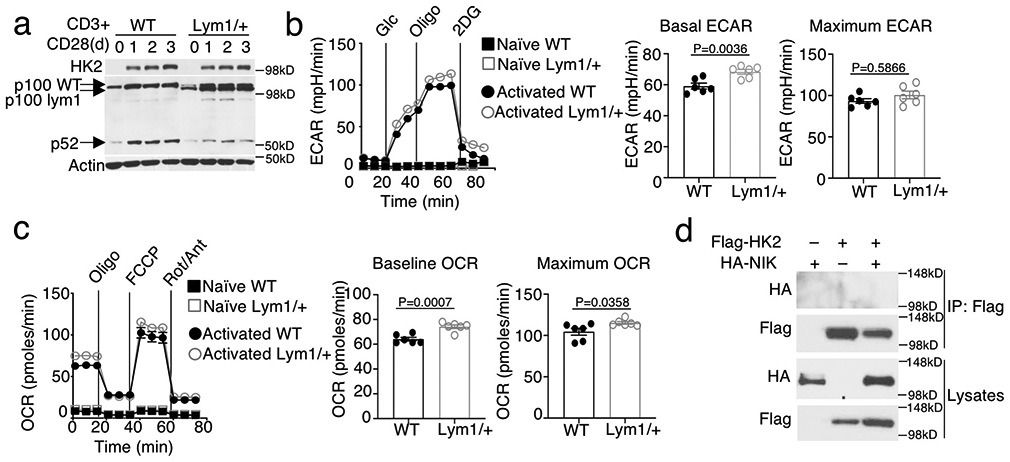
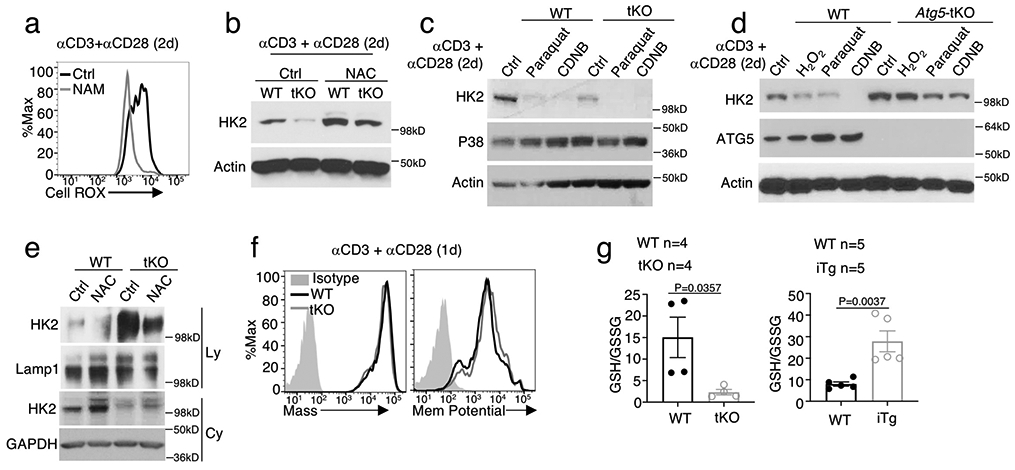
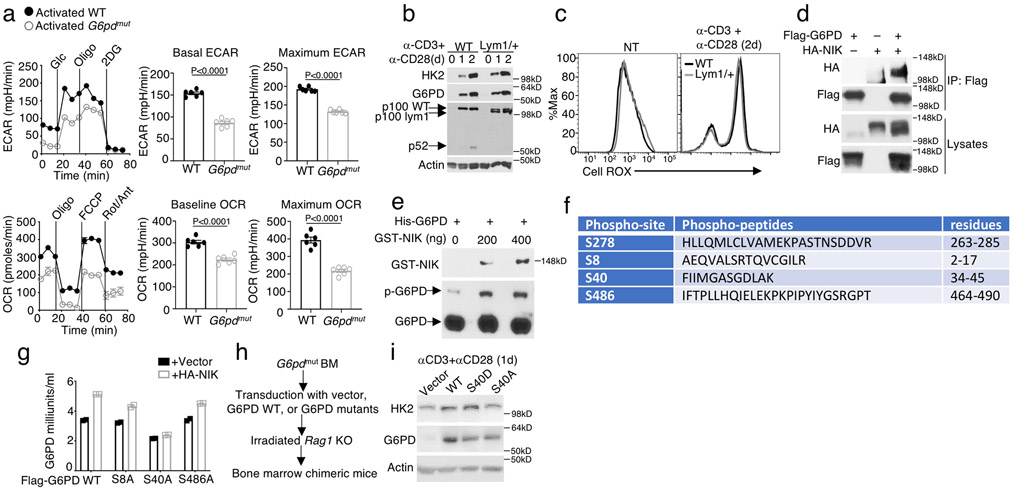
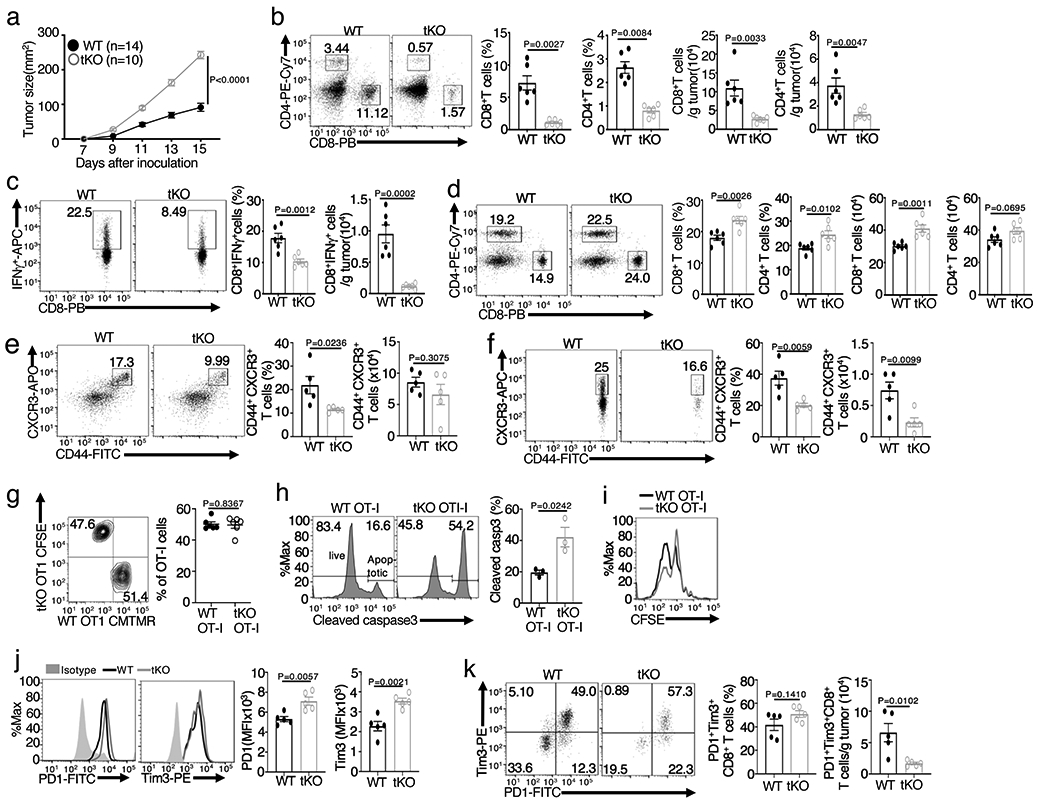
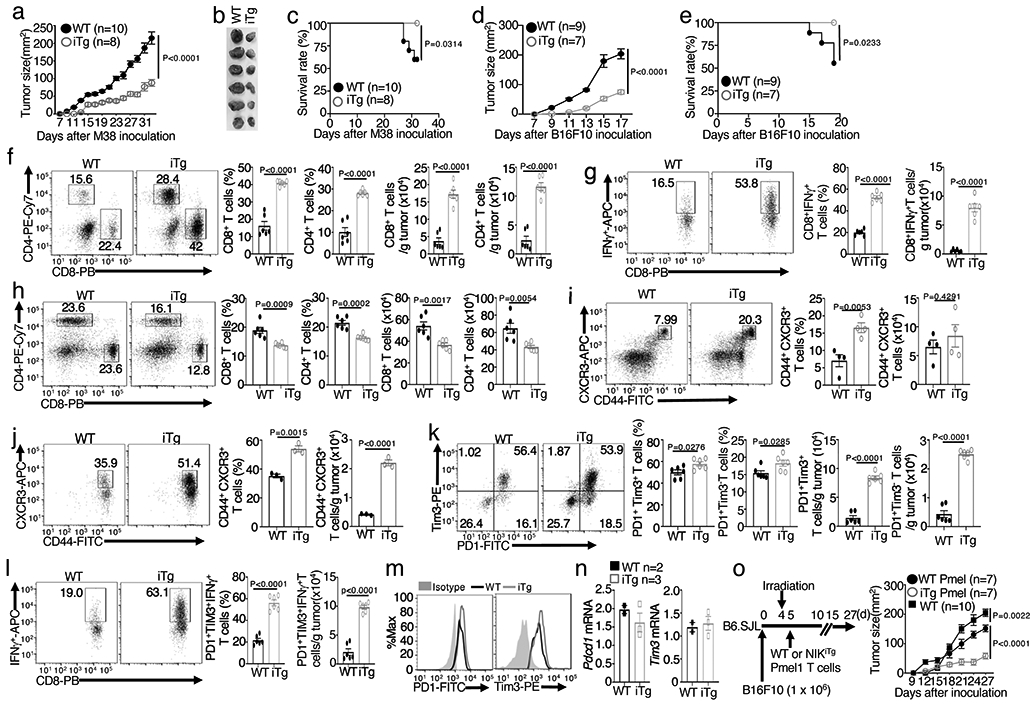
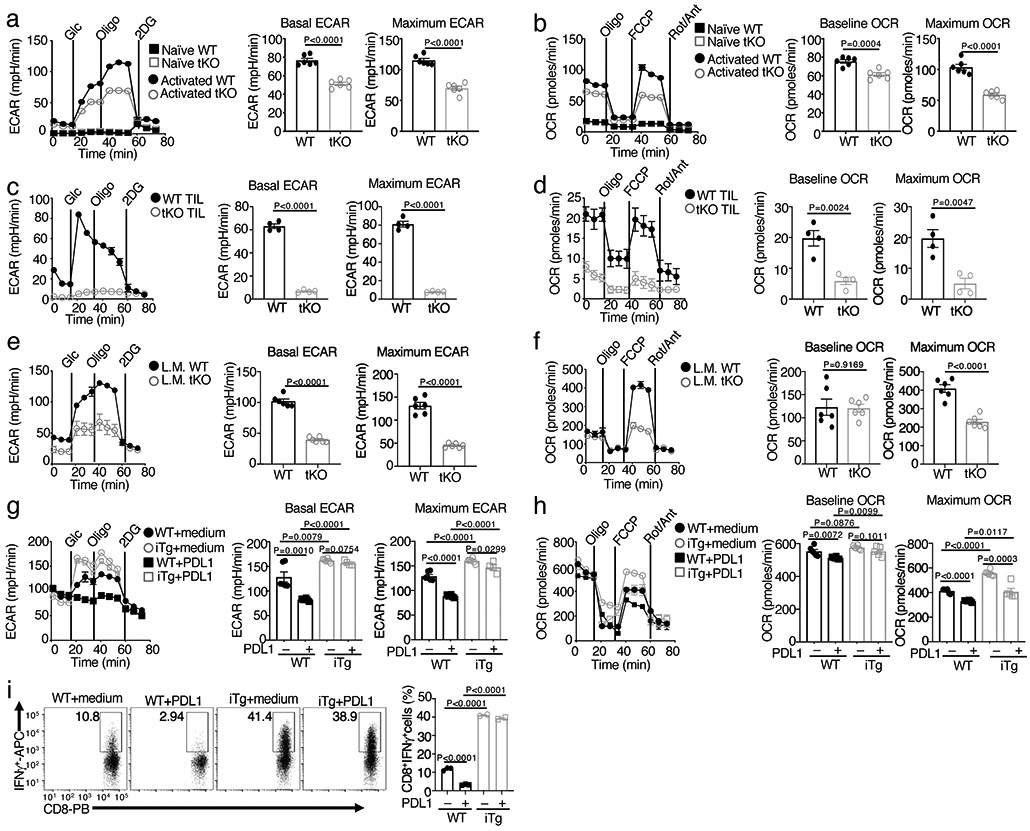
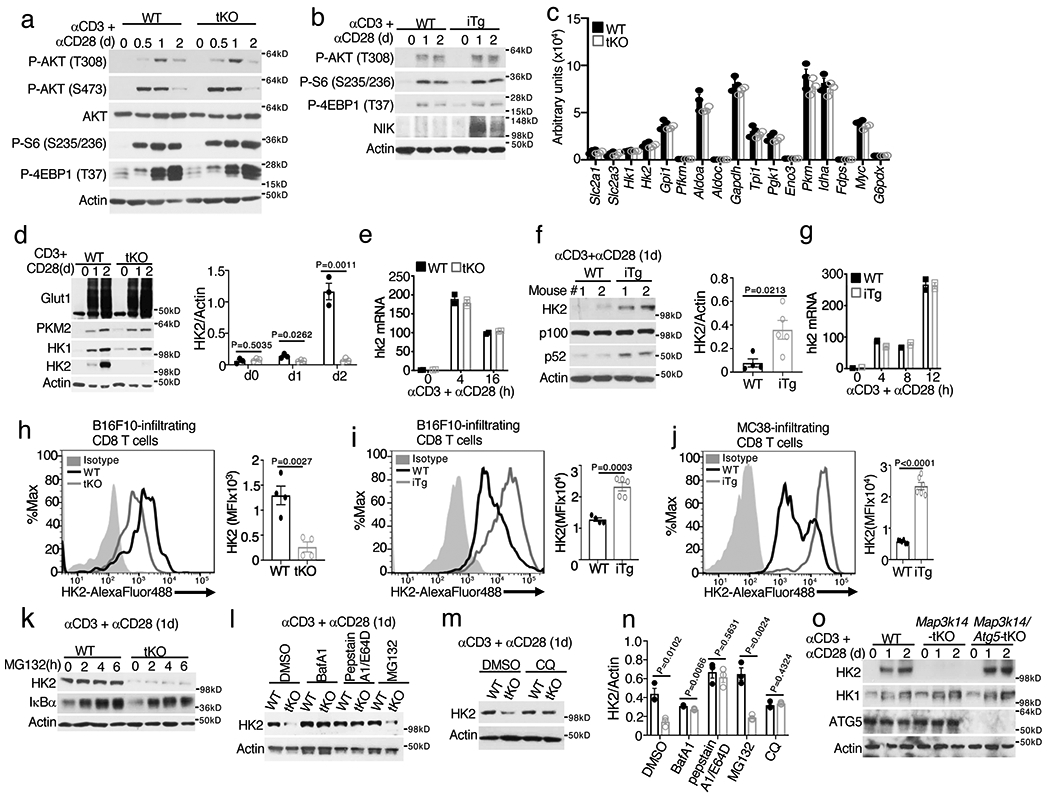
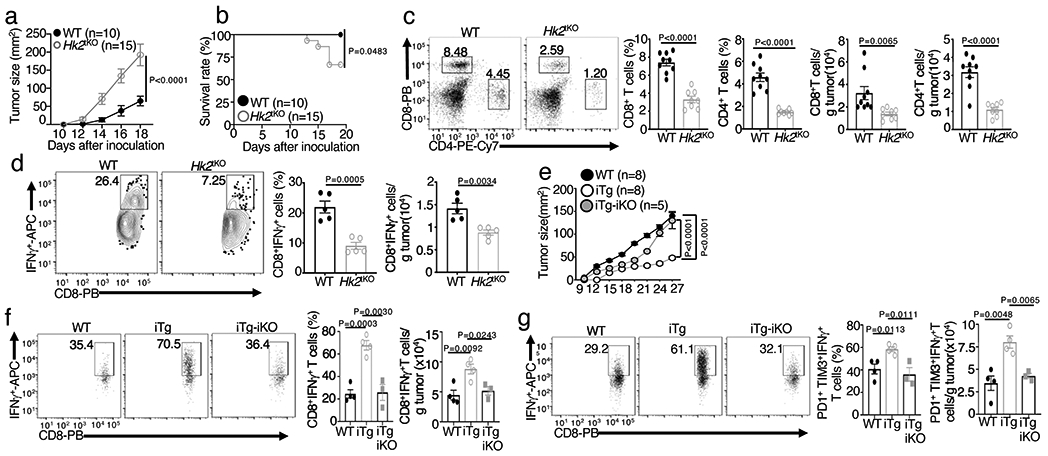
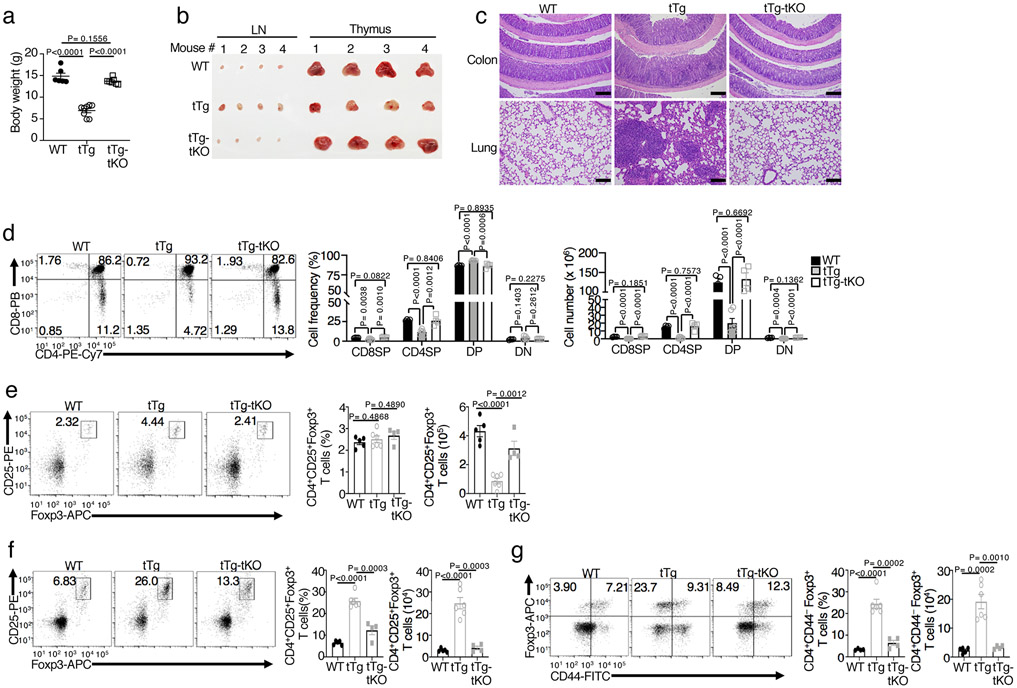
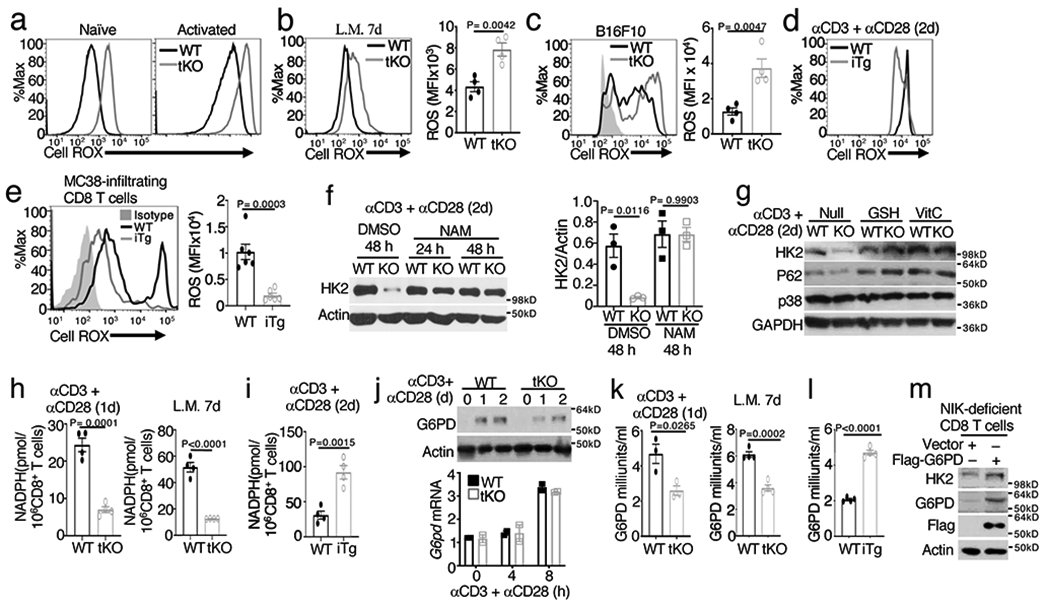
Metabolic reprograming toward aerobic glycolysis is a pivotal mechanism shaping immune responses. Here we show that deficiency in NF-κB-inducing kinase (NIK) impairs glycolysis induction, rendering CD8 effector T cells hypofunctional in the tumor microenvironment. Conversely, ectopic expression of NIK promotes CD8 T cell metabolism and effector function, thereby profoundly enhancing antitumor immunity and improving the efficacy of T cell adoptive therapy. NIK regulates T cell metabolism via a NF-κB-independent mechanism that involves stabilization of hexokinase 2 (HK2), a rate-limiting enzyme of the glycolytic pathway. NIK prevents autophagic degradation of HK2 through controlling cellular reactive oxygen species levels, which in turn involves modulation of glucose-6-phosphate dehydrogenase (G6PD), an enzyme that mediates production of the antioxidant NADPH. We show that the G6PD-NADPH redox system is important for HK2 stability and metabolism in activated T cells. These findings establish NIK as a pivotal regulator of T cell metabolism and highlight a post-translational mechanism of metabolic regulation.
代谢重编程向有氧糖酵解是塑造免疫反应的关键机制。在这里,我们表明,NF-κB 诱导激酶(NIK)的缺乏会损害糖酵解的诱导,使肿瘤微环境中的 CD8 效应 T 细胞功能低下。相反,NIK 的异位表达促进了 CD8 T 细胞的代谢和效应功能,从而显著增强了抗肿瘤免疫,并提高了 T 细胞过继治疗的疗效。NIK 通过一种不依赖 NF-κB 的机制来调节 T 细胞的代谢,该机制涉及到己糖激酶 2(HK2)的稳定,HK2 是糖酵解途径的限速酶。NIK 通过控制细胞内活性氧水平来防止 HK2 的自噬降解,这反过来又涉及葡萄糖-6-磷酸脱氢酶(G6PD)的调节,G6PD 是一种介导抗氧化剂 NADPH 产生的酶。我们表明,G6PD-NADPH 氧化还原系统对于激活的 T 细胞中 HK2 的稳定性和代谢是重要的。这些发现确立了 NIK 作为 T 细胞代谢的关键调节剂,并强调了代谢调节的一种翻译后机制。