Developmental Biology Unit, EMBL, Meyerhofstrasse 1, Heidelberg, 69117, Germany.
(Epi)genomics of Animal Development Unit, Department of Developmental and Stem Cell Biology, Institut Pasteur, 75015, Paris, France.
Nat Commun. 2021 Jan 19;12(1):439. doi: 10.1038/s41467-020-20714-y.
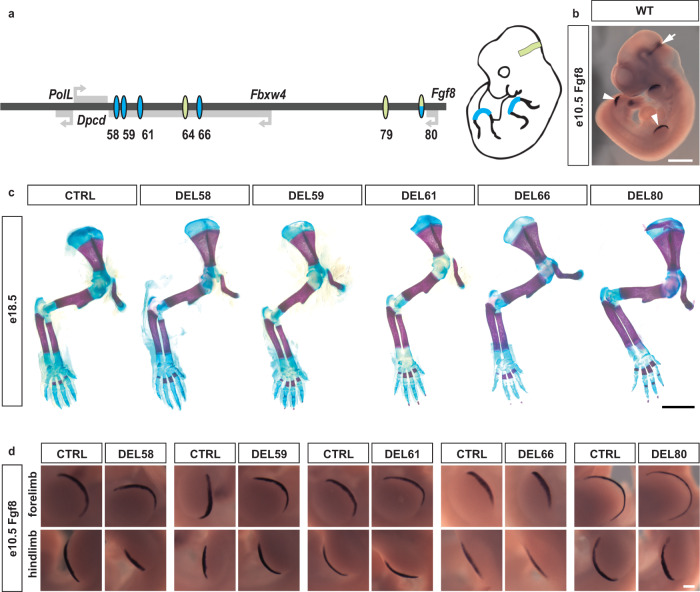
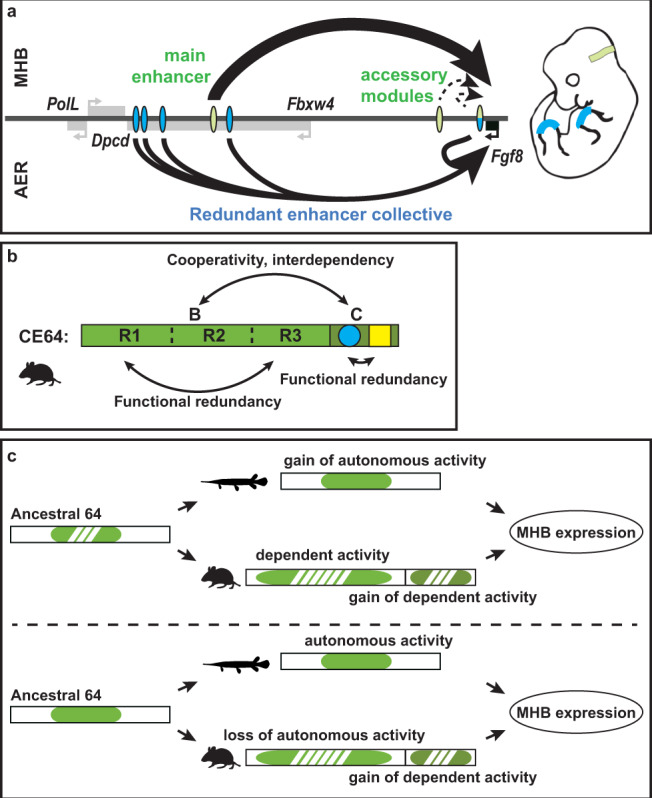
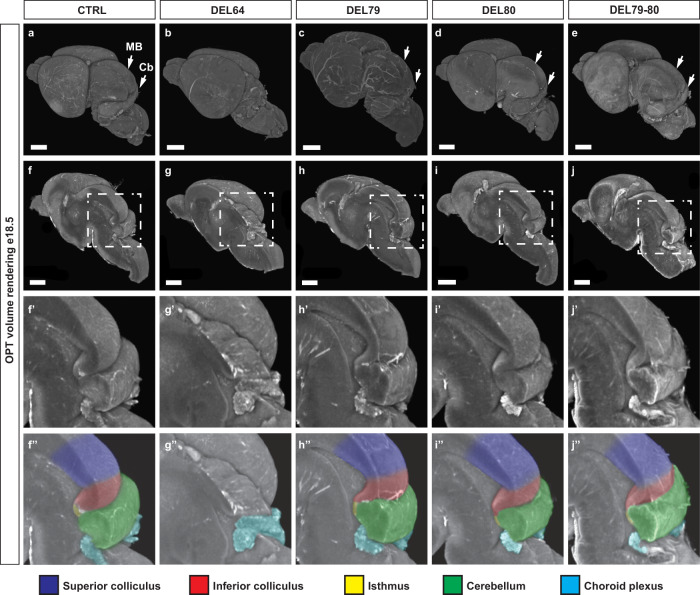
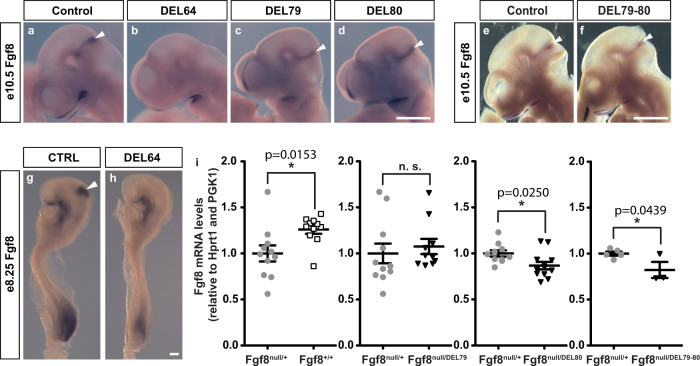
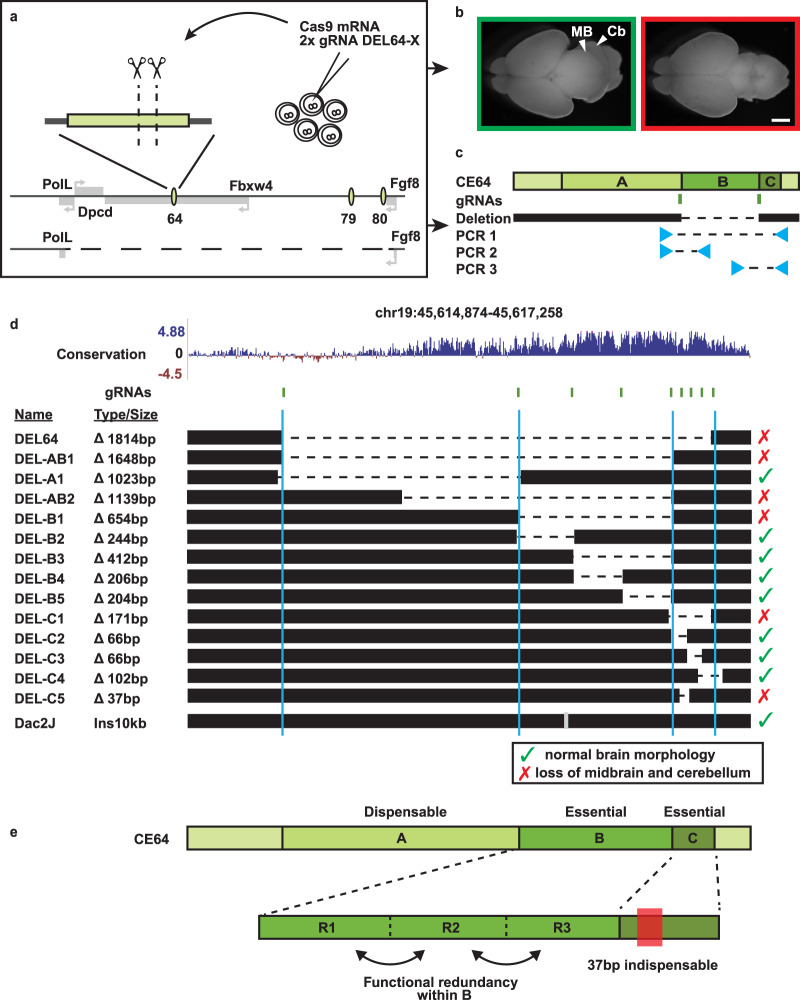
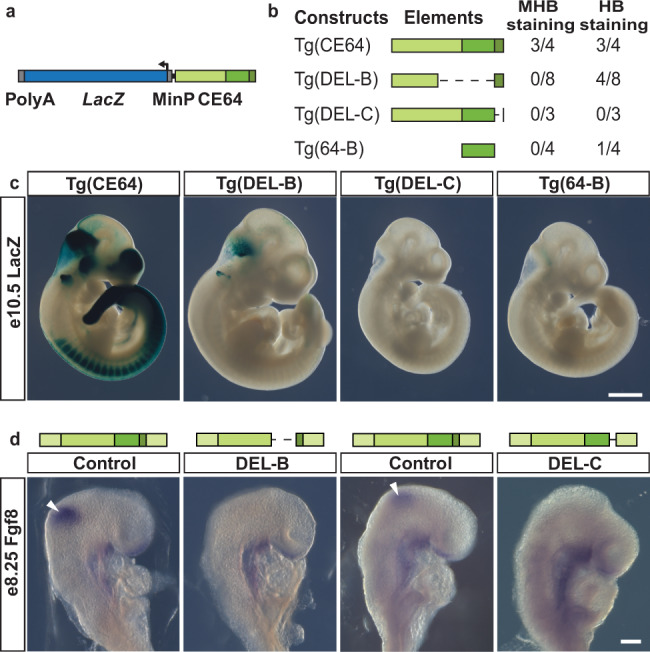
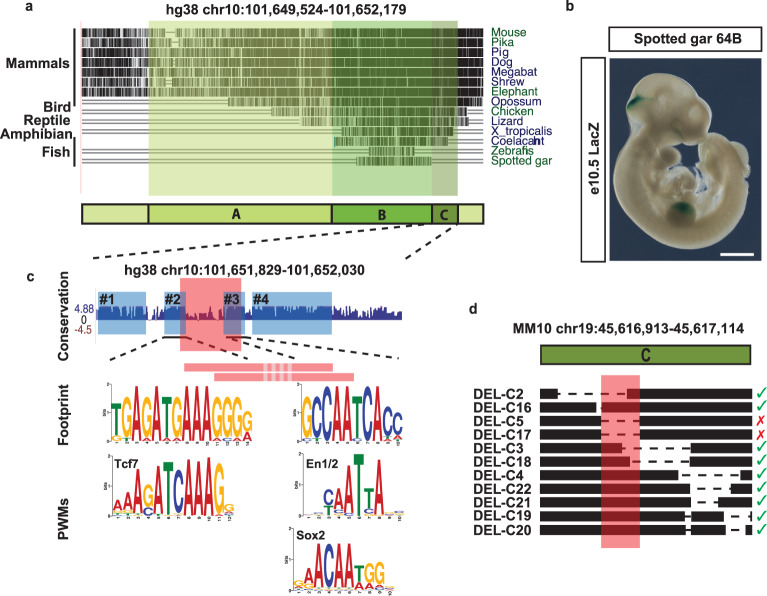
Developmental genes are often regulated by multiple elements with overlapping activity. Yet, in most cases, the relative function of those elements and their contribution to endogenous gene expression remain poorly characterized. An example of this phenomenon is that distinct sets of enhancers have been proposed to direct Fgf8 in the limb apical ectodermal ridge and the midbrain-hindbrain boundary. Using in vivo CRISPR/Cas9 genome engineering, we functionally dissect this complex regulatory ensemble and demonstrate two distinct regulatory logics. In the apical ectodermal ridge, the control of Fgf8 expression appears distributed between different enhancers. In contrast, we find that in the midbrain-hindbrain boundary, one of the three active enhancers is essential while the other two are dispensable. We further dissect the essential midbrain-hindbrain boundary enhancer to reveal that it is also composed by a mixture of essential and dispensable modules. Cross-species transgenic analysis of this enhancer suggests that its composition may have changed in the vertebrate lineage.
发育基因通常受多个具有重叠活性的元件调控。然而,在大多数情况下,这些元件的相对功能及其对内源性基因表达的贡献仍未得到很好的描述。这种现象的一个例子是,已经提出了不同的增强子集合来指导肢端顶外胚层嵴和中脑-后脑边界处的 Fgf8。我们使用体内 CRISPR/Cas9 基因组工程,从功能上对这个复杂的调控组件进行了剖析,并证明了两种不同的调控逻辑。在顶外胚层嵴中,Fgf8 表达的控制似乎分布在不同的增强子之间。相比之下,我们发现,在中脑-后脑边界处,三个活跃的增强子中有一个是必需的,而另外两个是可有可无的。我们进一步剖析必需的中脑-后脑边界增强子,揭示其也由必需和可有可无的模块组成。对该增强子的跨物种转基因分析表明,其组成在脊椎动物谱系中可能已经发生了变化。