Laboratory for Phyloinformatics, RIKEN Center for Biosystems Dynamics Research (BDR), Kobe, Japan.
Laboratory for Bioinformatics Research, RIKEN BDR, Wako City, Japan.
Elife. 2021 Feb 9;10:e62865. doi: 10.7554/eLife.62865.
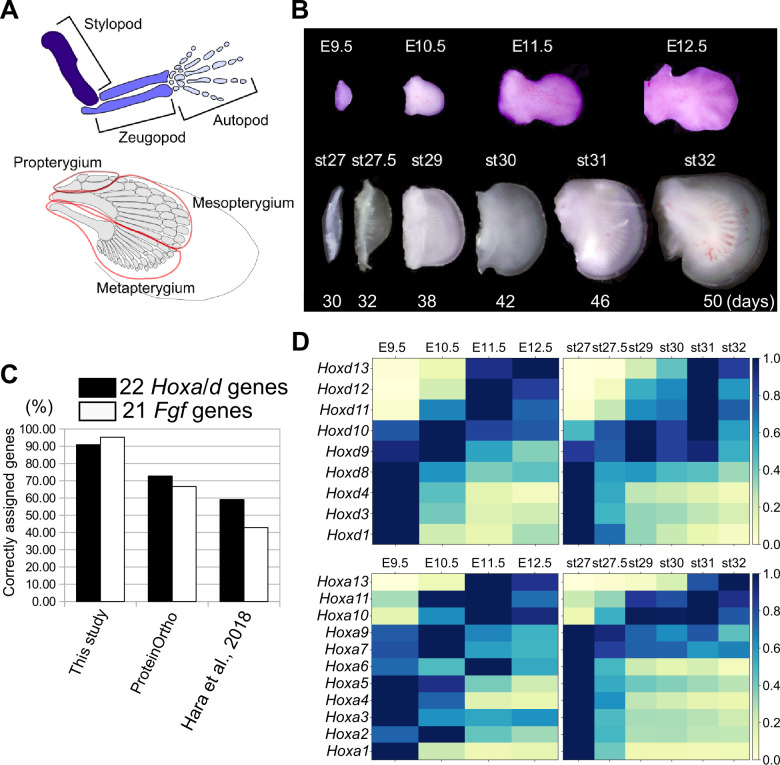
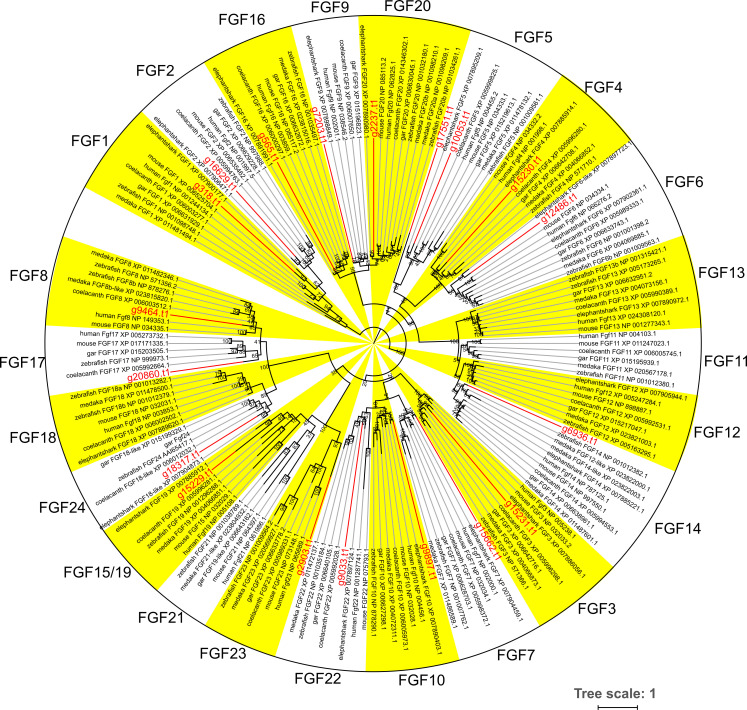
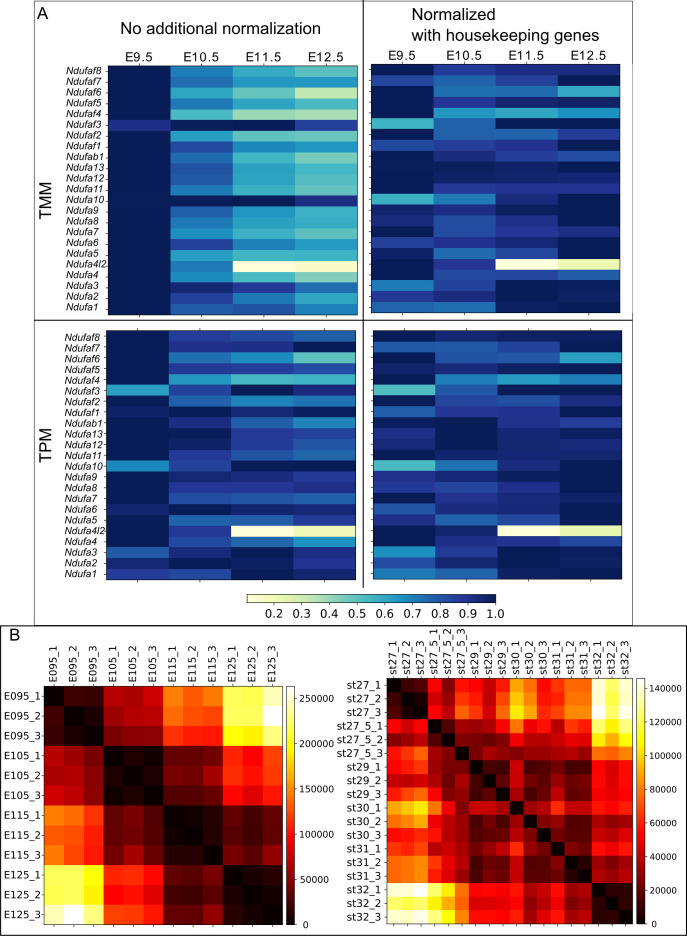
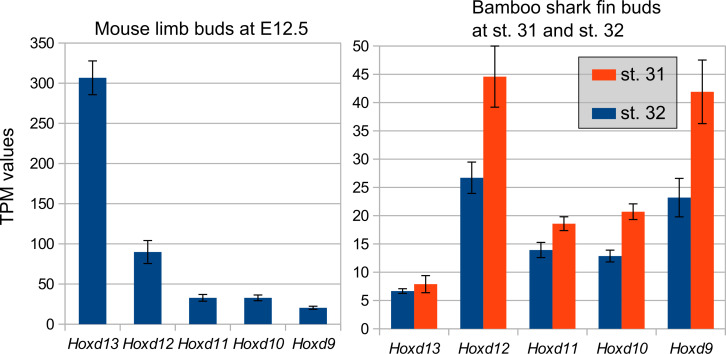
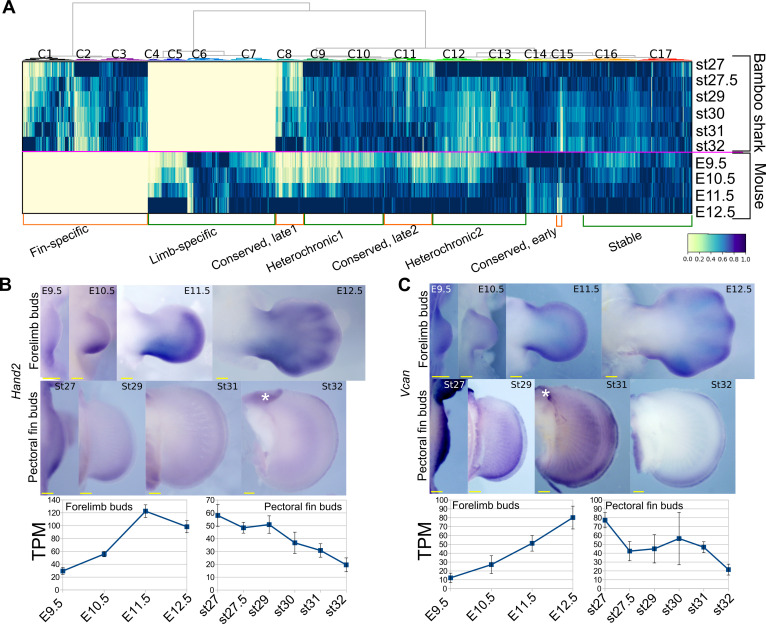
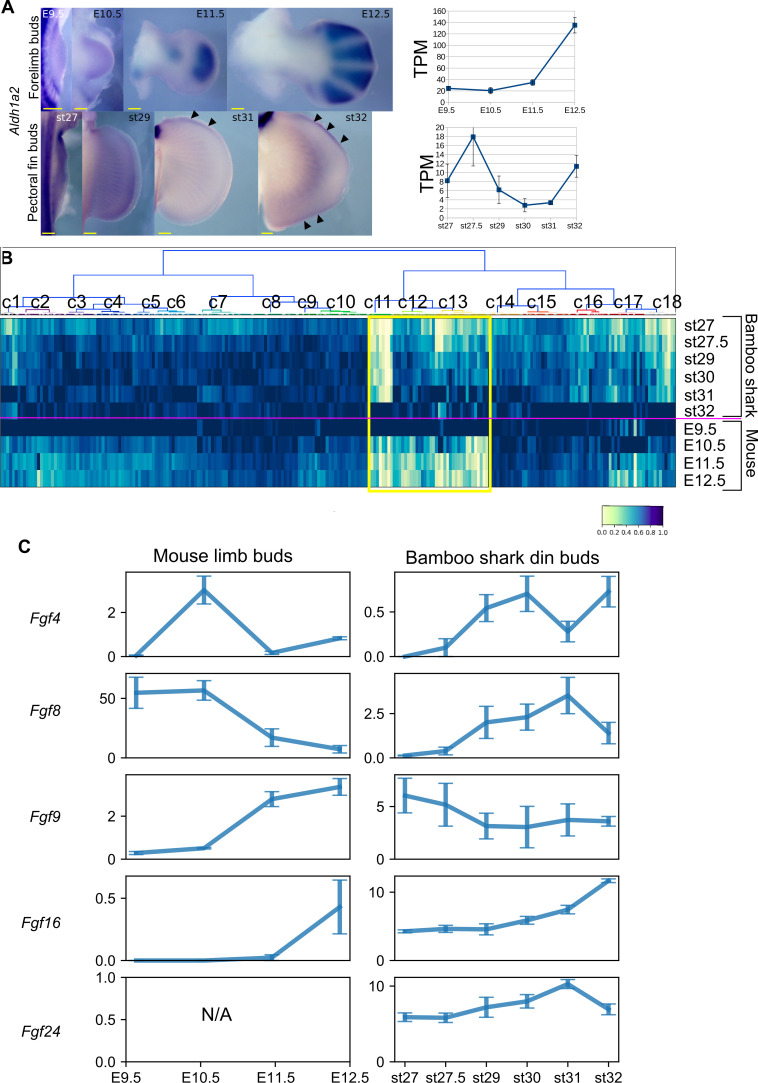
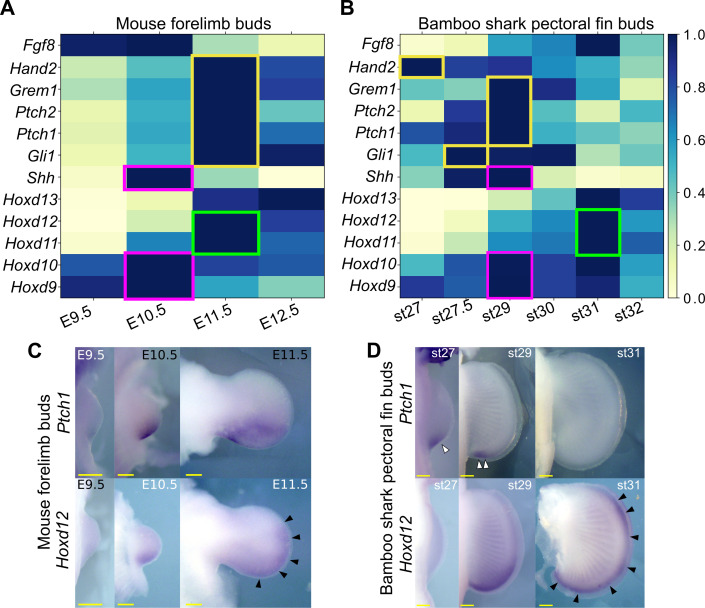
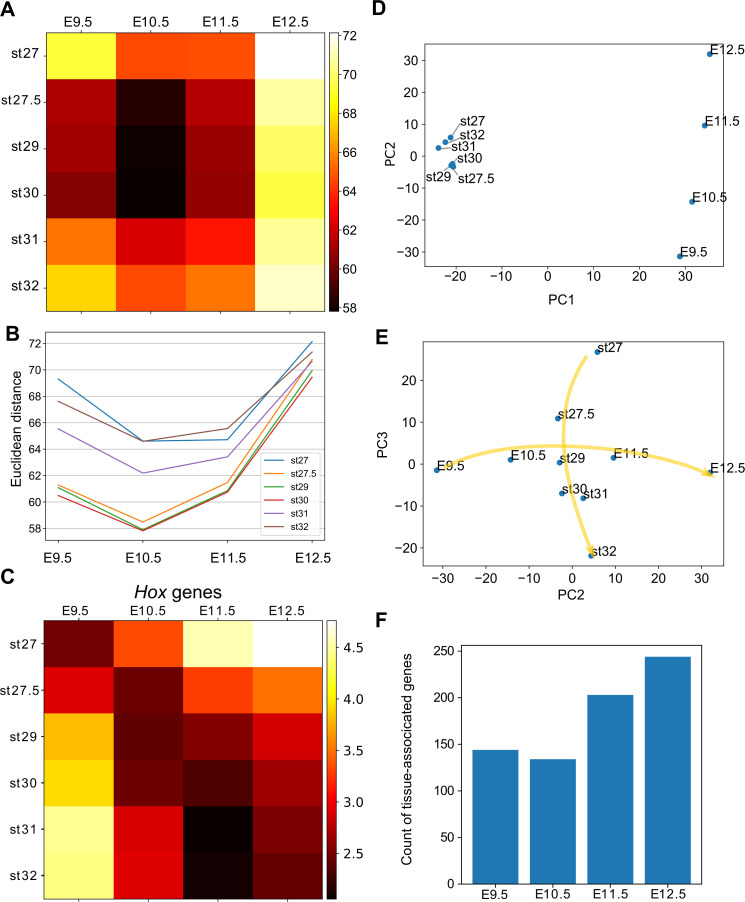
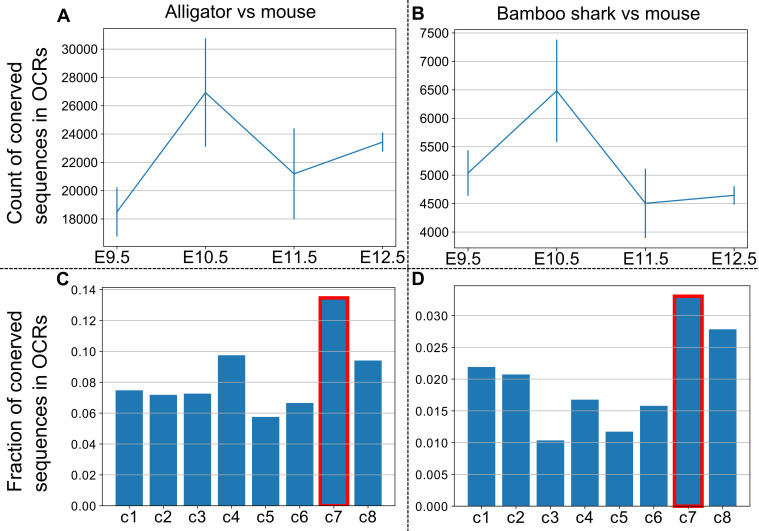
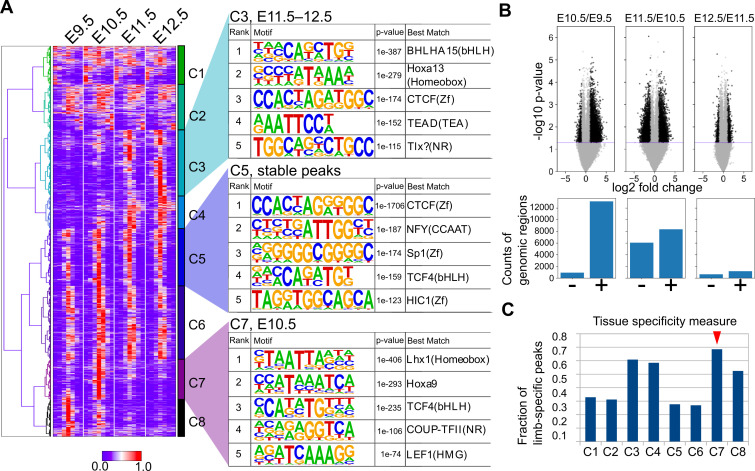
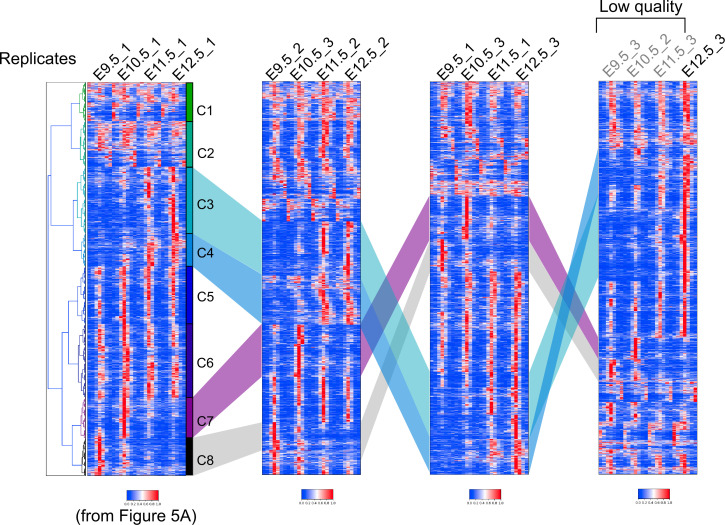
How genetic changes are linked to morphological novelties and developmental constraints remains elusive. Here, we investigate genetic apparatuses that distinguish fish fins from tetrapod limbs by analyzing transcriptomes and open-chromatin regions (OCRs). Specifically, we compared mouse forelimb buds with the pectoral fin buds of an elasmobranch, the brown-banded bamboo shark (). A transcriptomic comparison with an accurate orthology map revealed both a mass heterochrony and hourglass-shaped conservation of gene expression between fins and limbs. Furthermore, open-chromatin analysis suggested that access to conserved regulatory sequences is transiently increased during mid-stage limb development. During this stage, stage-specific and tissue-specific OCRs were also enriched. Together, early and late stages of fin/limb development are more permissive to mutations than middle stages, which may have contributed to major morphological changes during the fin-to-limb evolution. We hypothesize that the middle stages are constrained by regulatory complexity that results from dynamic and tissue-specific transcriptional controls.
遗传变化如何与形态创新和发育限制相关仍然难以捉摸。在这里,我们通过分析转录组和开放染色质区域(OCR)来研究区分鱼类鳍和四足动物肢体的遗传装置。具体来说,我们比较了鼠前肢芽和软骨鱼纲的胸鳍芽,即棕带竹鲨(Chiloscyllium plagiosum)。与准确的同线性图谱进行转录组比较显示,鳍和肢体之间存在大规模的异时性和沙漏形的基因表达保守性。此外,开放染色质分析表明,在肢体发育的中期,保守调控序列的可及性暂时增加。在这个阶段,阶段特异性和组织特异性 OCR 也被富集。总的来说,鳍/肢发育的早期和晚期比中期更允许突变,这可能导致鳍到肢进化过程中的主要形态变化。我们假设,中间阶段受到来自动态和组织特异性转录控制的调节复杂性的限制。