Department of Physics, Technical University of Munich, Garching, Germany.
Molecular Biology Division, Biomedical Center, Faculty of Medicine, Ludwig-Maximilians-Universität München, Planegg-Martinsried, Germany.
Elife. 2021 Mar 5;10:e58394. doi: 10.7554/eLife.58394.
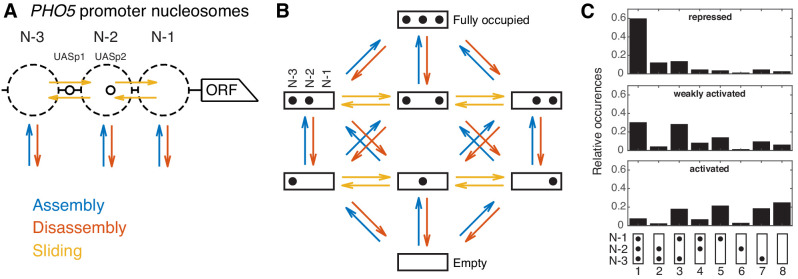
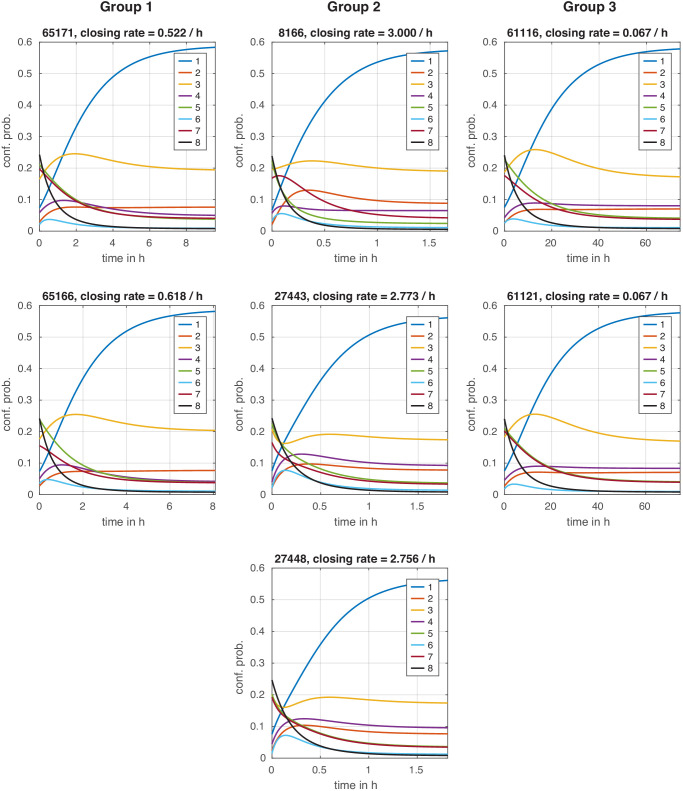
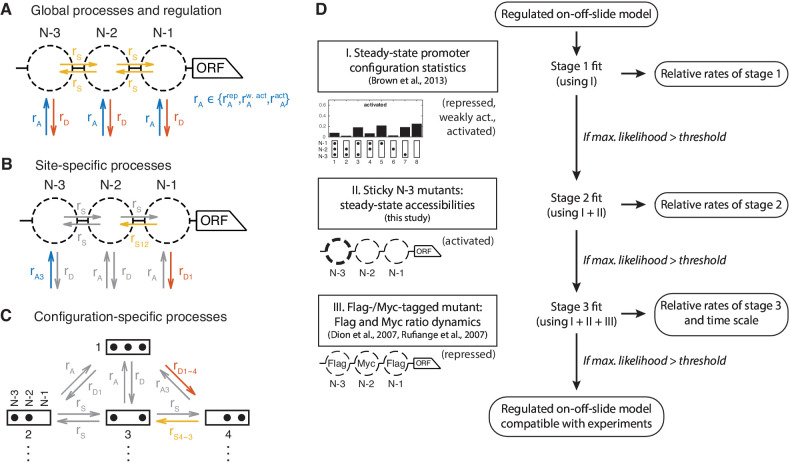
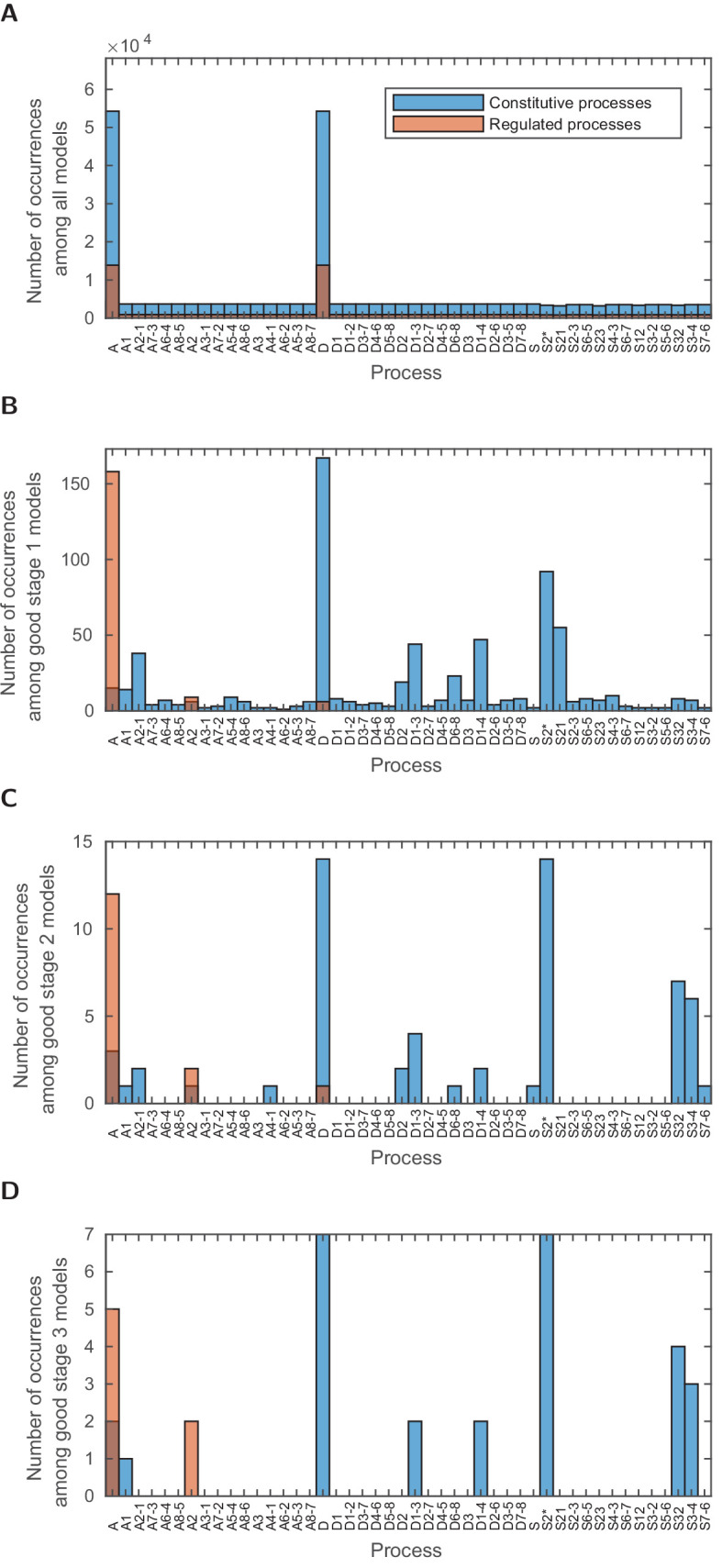
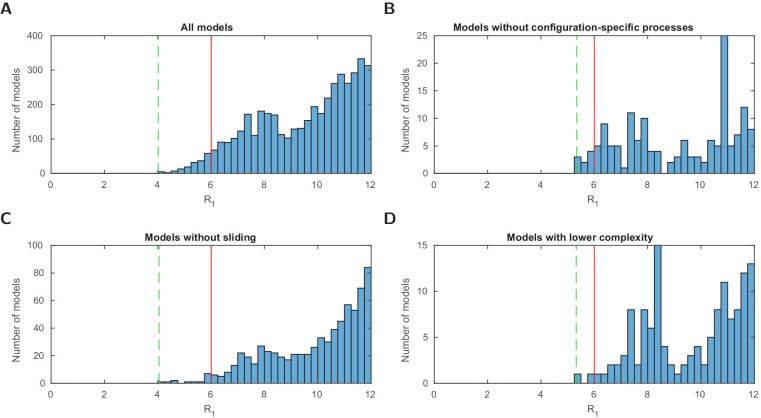
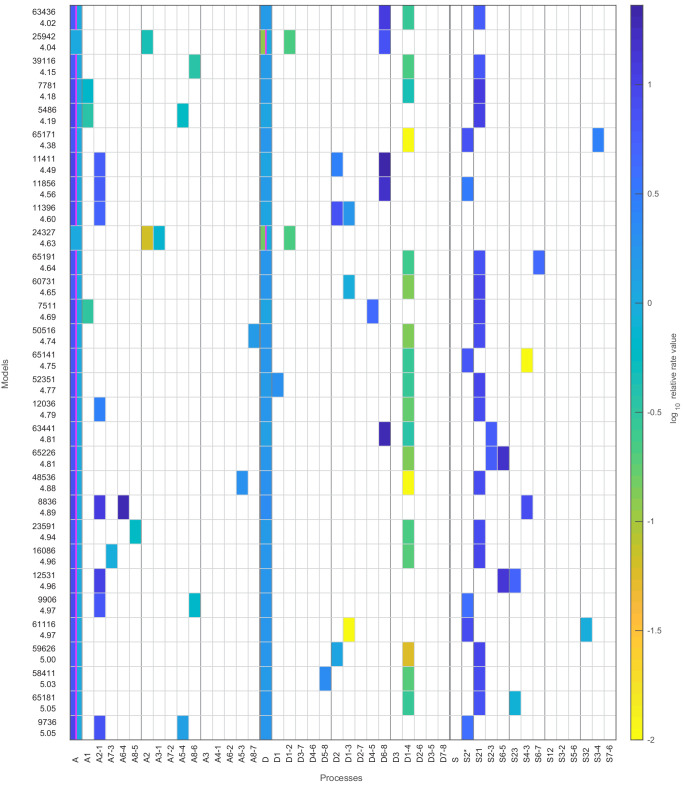
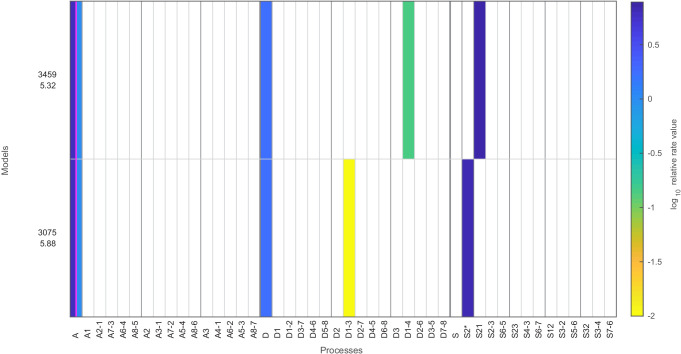
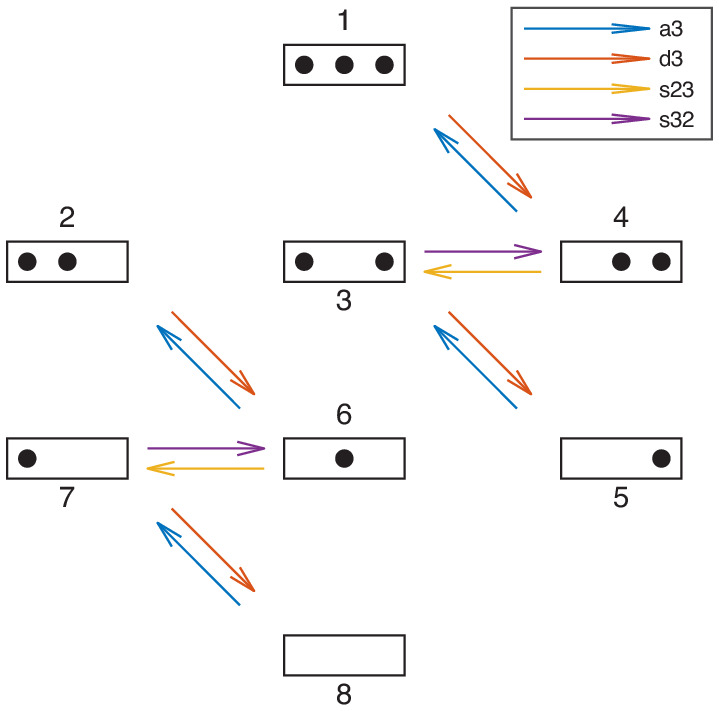
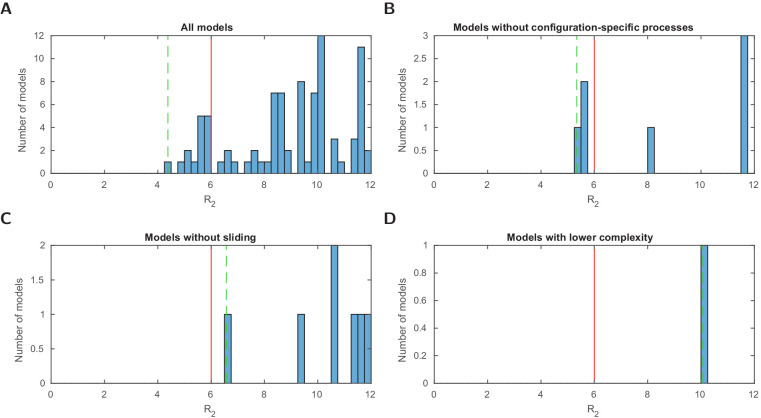
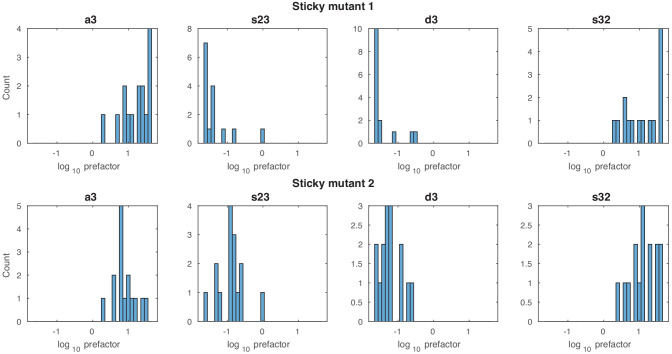
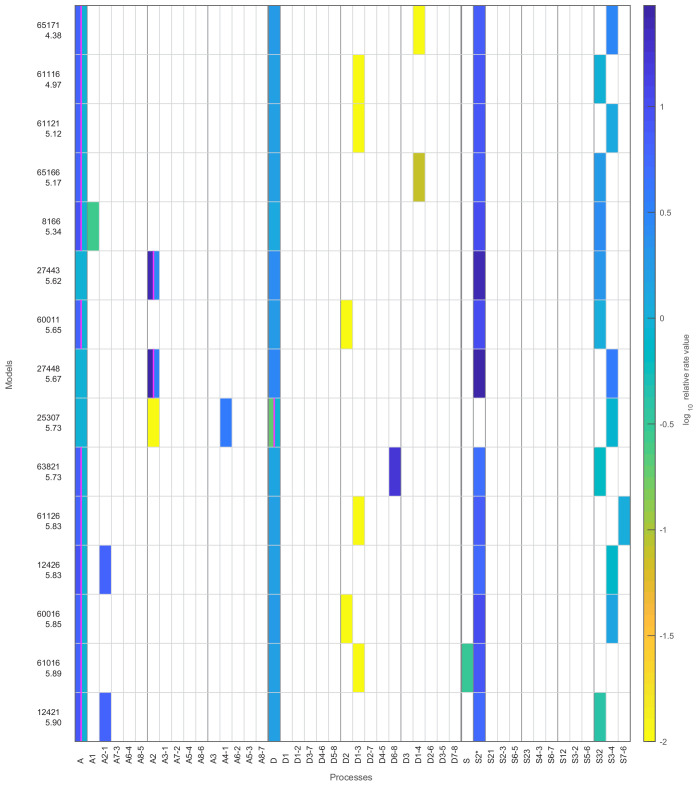
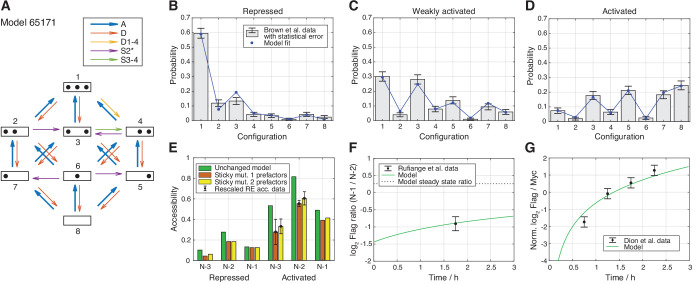
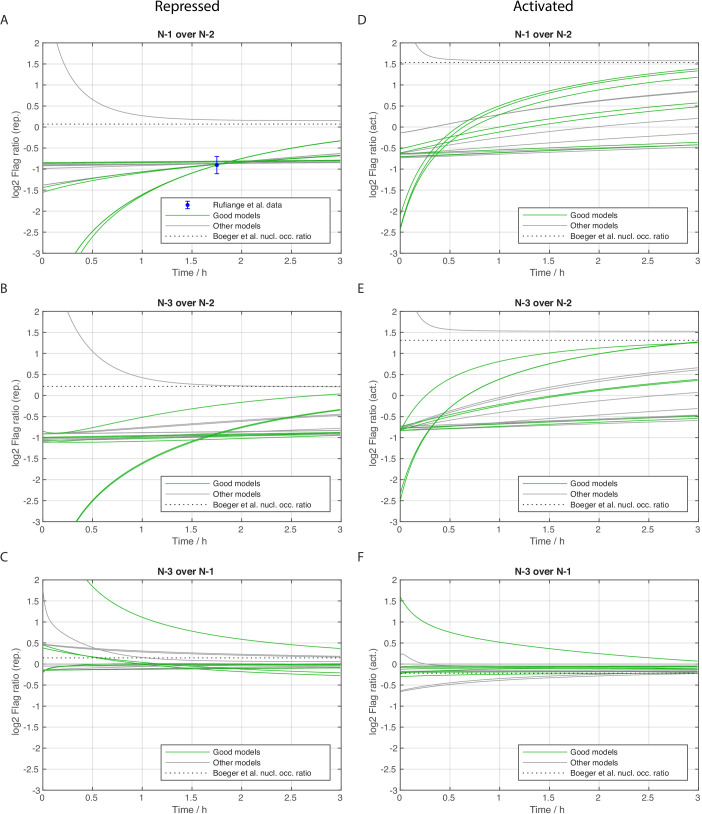
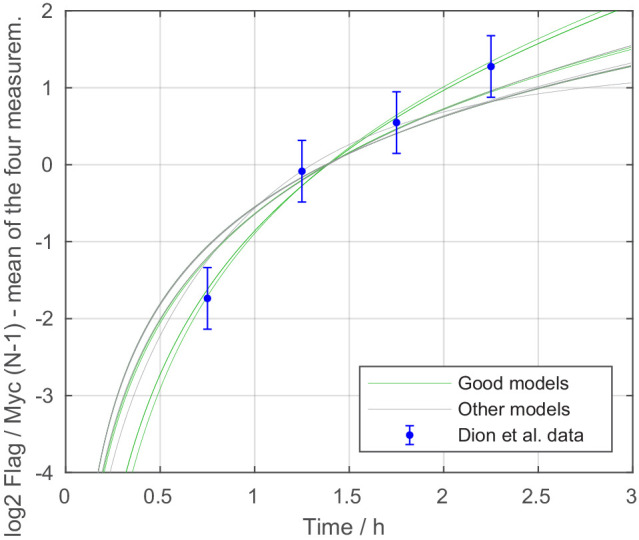
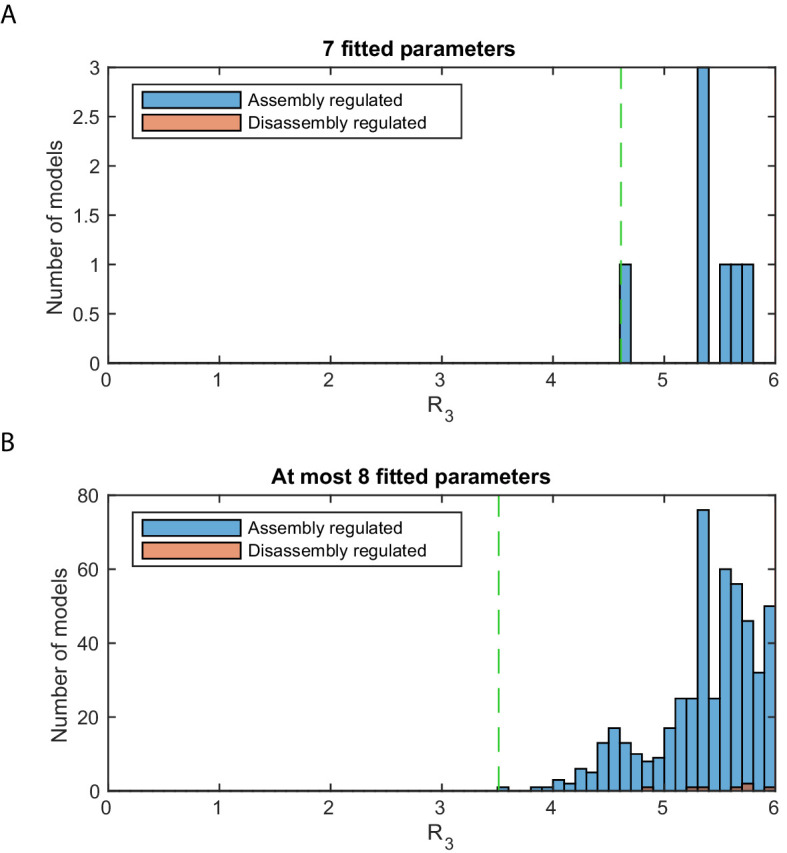
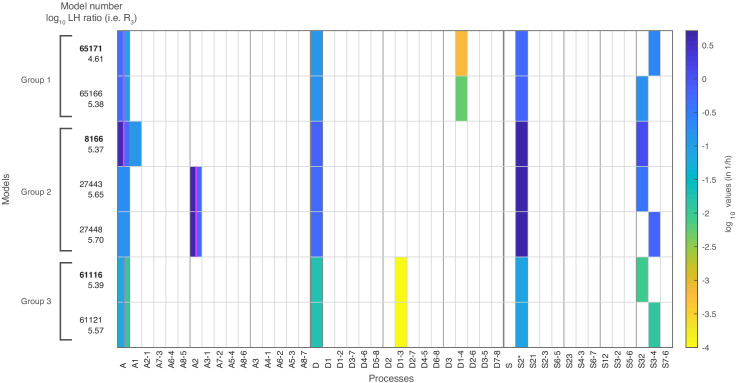
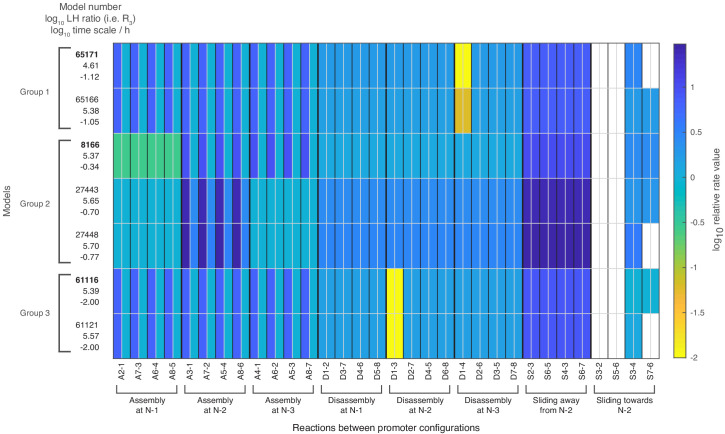
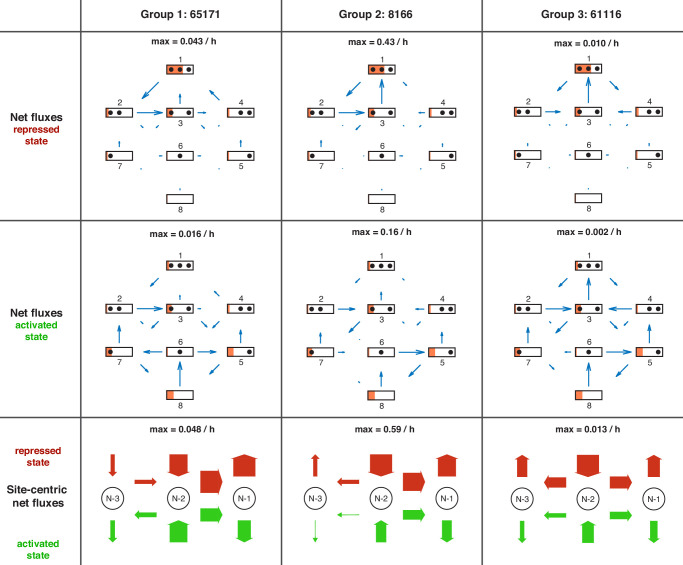
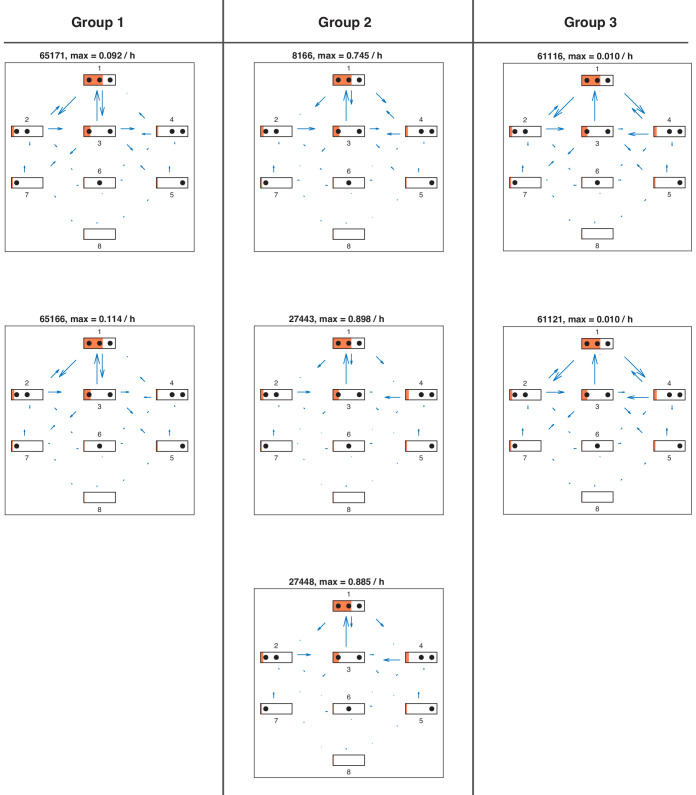
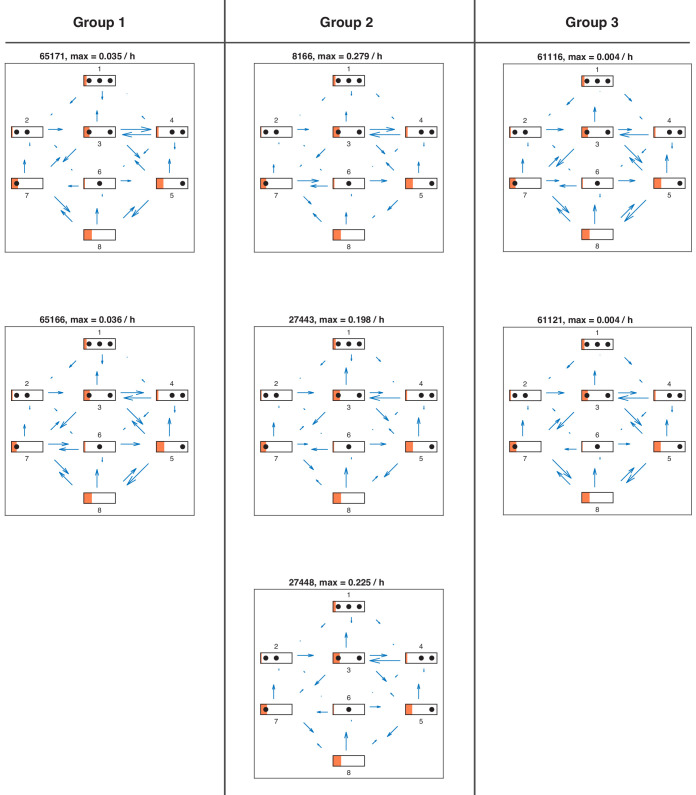
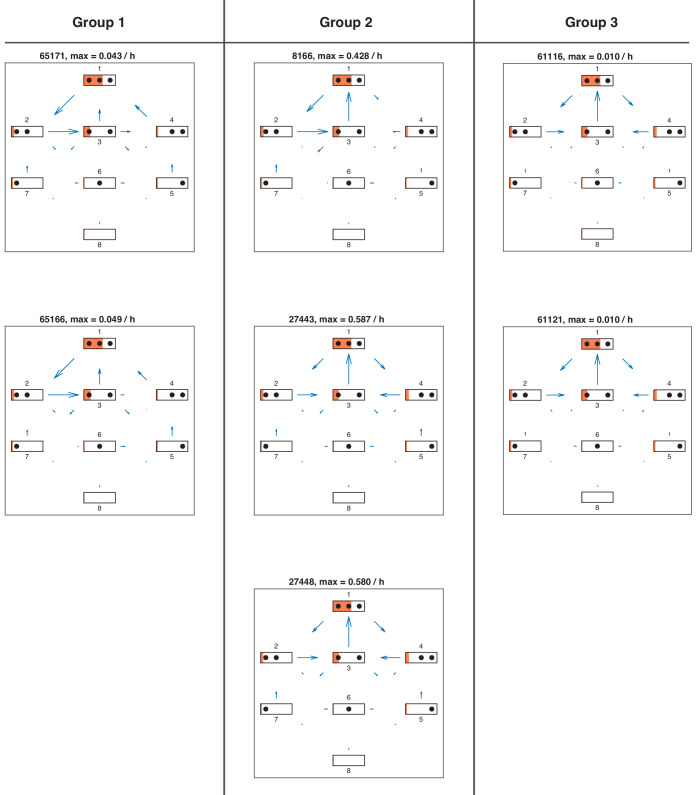
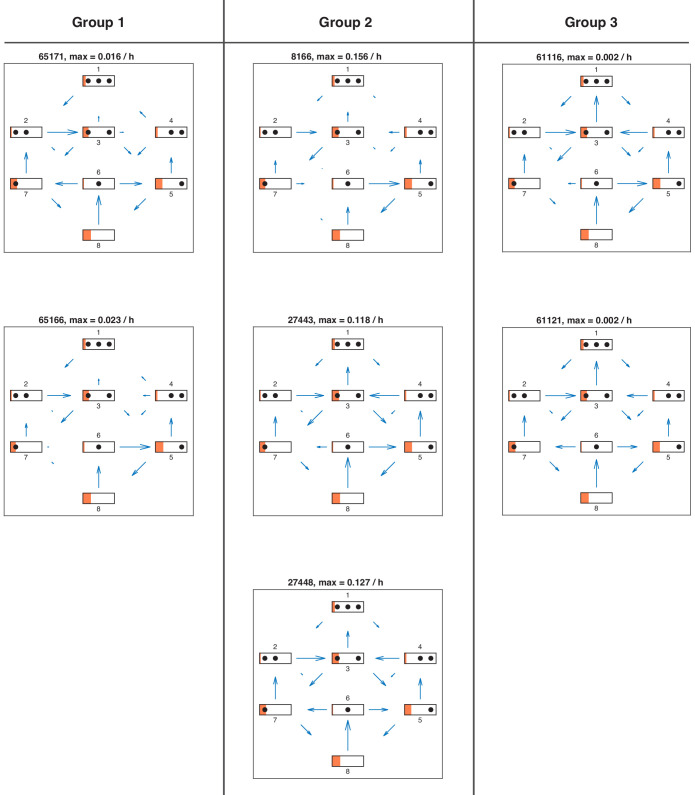
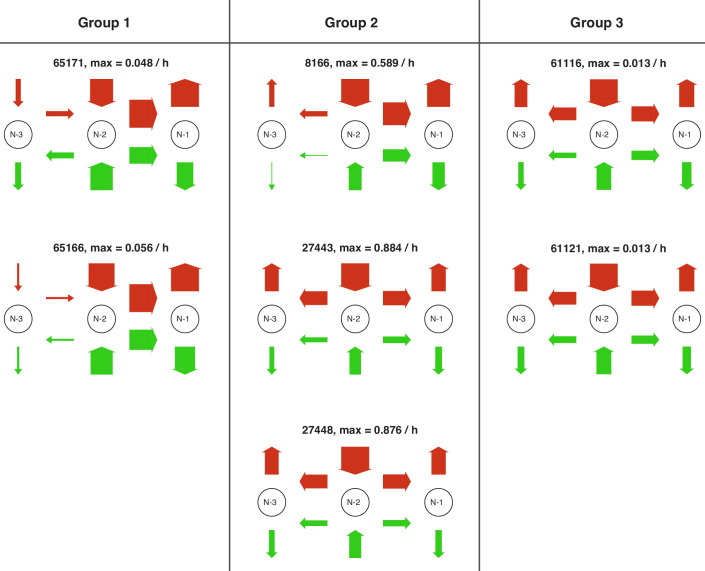
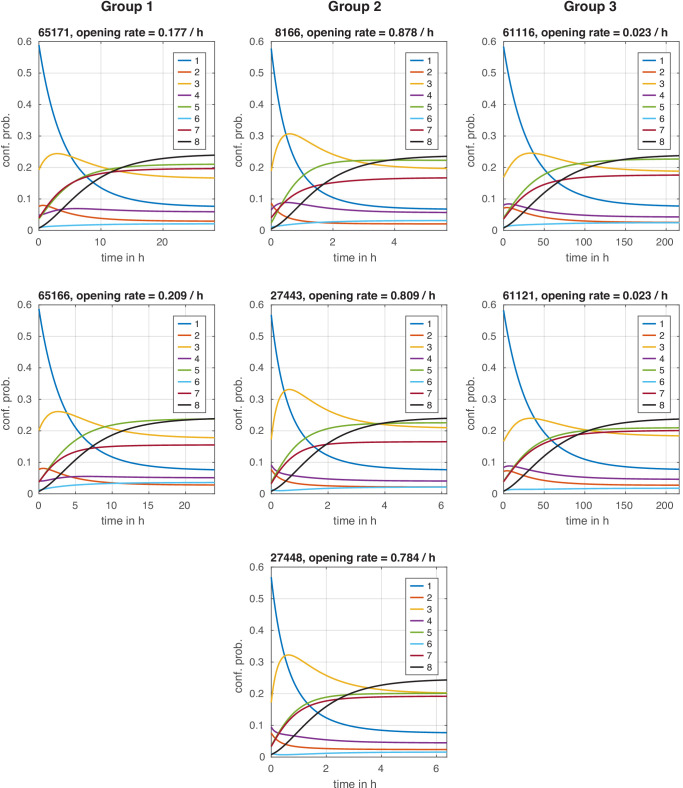
Chromatin dynamics are mediated by remodeling enzymes and play crucial roles in gene regulation, as established in a paradigmatic model, the promoter. However, effective nucleosome dynamics, that is, trajectories of promoter nucleosome configurations, remain elusive. Here, we infer such dynamics from the integration of published single-molecule data capturing multi-nucleosome configurations for repressed to fully active promoter states with other existing histone turnover and new chromatin accessibility data. We devised and systematically investigated a new class of 'regulated on-off-slide' models simulating global and local nucleosome (dis)assembly and sliding. Only seven of 68,145 models agreed well with all data. All seven models involve sliding and the known central role of the N-2 nucleosome, but regulate promoter state transitions by modulating just one assembly rather than disassembly process. This is consistent with but challenges common interpretations of previous observations at the promoter and suggests chromatin opening by binding competition.
染色质动力学由重塑酶介导,并在基因调控中发挥关键作用,这在一个典范模型——启动子中得到了证实。然而,有效的核小体动力学,即启动子核小体构象的轨迹,仍然难以捉摸。在这里,我们从整合已发表的单分子数据中推断出这种动力学,这些数据捕获了从被抑制到完全激活的启动子状态的多核小体构象,同时还整合了其他现有的组蛋白周转率和新的染色质可及性数据。我们设计并系统地研究了一类新的“调节开-关滑”模型,模拟了全局和局部核小体(解)组装和滑动。在 68145 个模型中,只有 7 个与所有数据吻合得很好。这 7 个模型都涉及滑动和已知的 N-2 核小体的核心作用,但通过调节一个组装过程而不是解组装过程来调节启动子状态的转变。这与之前在启动子上观察到的结果一致,但也对其提出了挑战,并表明染色质的打开是通过结合竞争实现的。