Immune Cell Biology Laboratory, Department of Biomedicine, University of Basel, University Hospital Basel, Basel, Switzerland.
Division of Experimental Virology, Department of Biomedicine - Haus Petersplatz, University of Basel, Basel, Switzerland.
Elife. 2021 Mar 8;10:e61869. doi: 10.7554/eLife.61869.
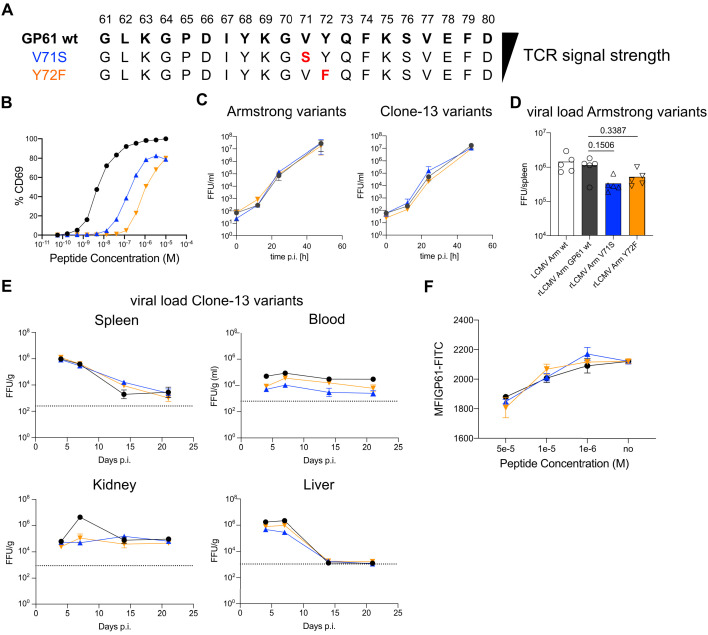
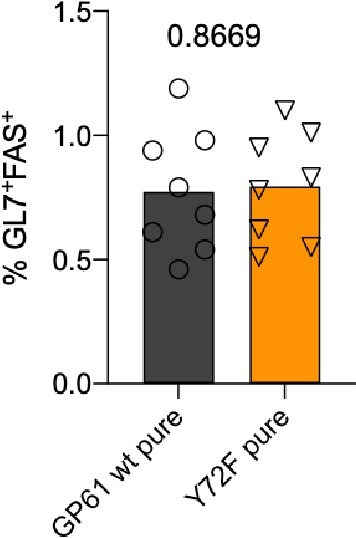
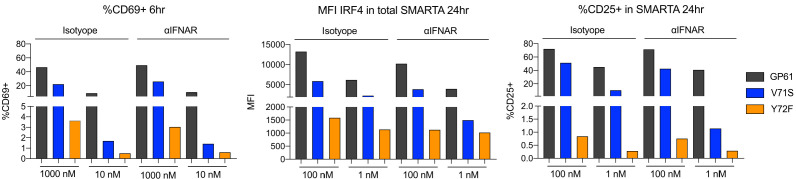
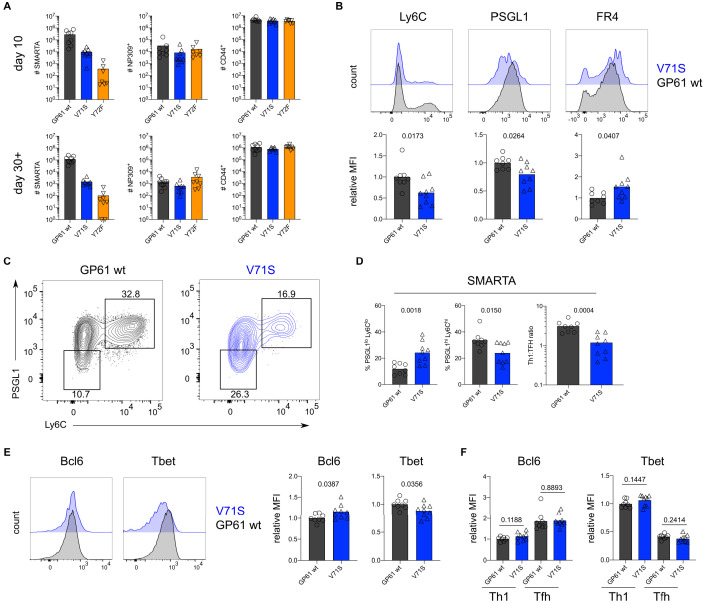
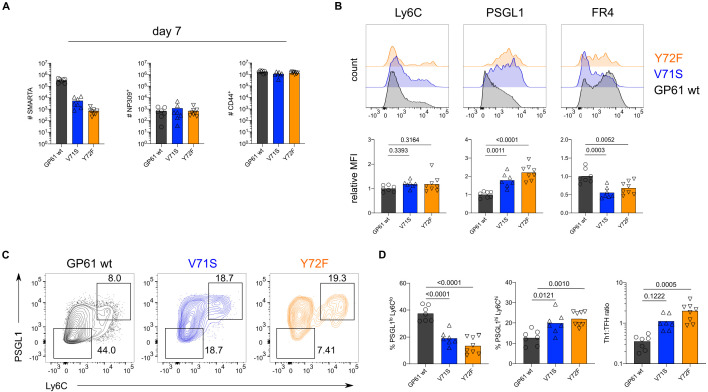
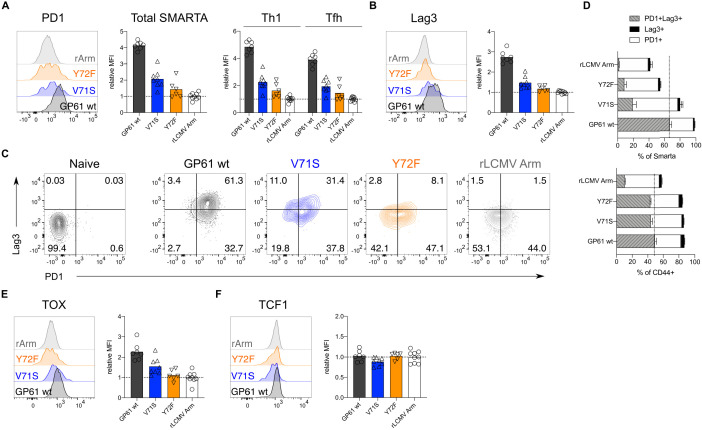
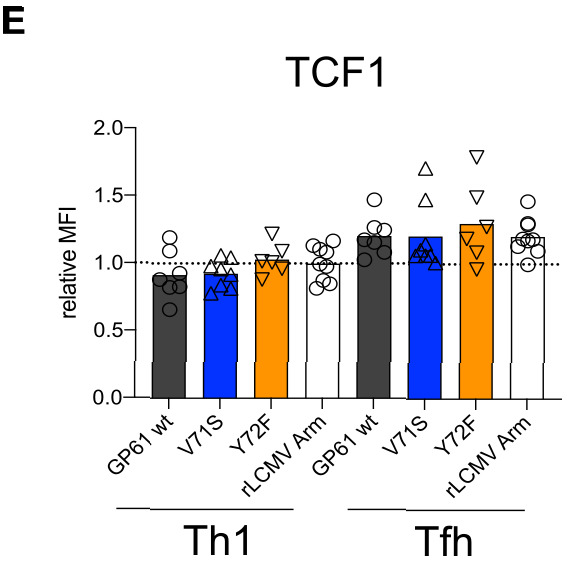
A hallmark of adaptive immunity is CD4 T cells' ability to differentiate into specialized effectors. A long-standing question is whether T cell receptor (TCR) signal strength can dominantly instruct the development of Th1 and T follicular helper (Tfh) cells across distinct infectious contexts. We characterized the differentiation of murine CD4 TCR transgenic T cells responding to altered peptide ligand lymphocytic choriomeningitis viruses (LCMV) derived from acute and chronic parental strains. We found that TCR signal strength exerts opposite and hierarchical effects on the balance of Th1 and Tfh cells responding to acute versus persistent infection. TCR signal strength correlates positively with Th1 generation during acute but negatively during chronic infection. Weakly activated T cells express lower levels of markers associated with chronic T cell stimulation and may resist functional inactivation. We anticipate that the panel of recombinant viruses described herein will be valuable for investigating a wide range of CD4 T cell responses.
适应性免疫的一个标志是 CD4 T 细胞将分化为专门的效应细胞的能力。一个长期存在的问题是,T 细胞受体(TCR)信号强度是否可以主导 Th1 和滤泡辅助 T 细胞(Tfh)细胞在不同感染环境下的发育。我们描述了对源自急性和慢性亲本株的改变肽配基淋巴细胞性脉络丛脑膜炎病毒(LCMV)的小鼠 CD4 TCR 转基因 T 细胞的分化。我们发现,TCR 信号强度对急性和持续性感染中 Th1 和 Tfh 细胞平衡的影响相反且具有层次结构。在急性感染期间,TCR 信号强度与 Th1 生成呈正相关,但在慢性感染期间呈负相关。弱激活的 T 细胞表达与慢性 T 细胞刺激相关的标志物水平较低,并且可能抵抗功能失活。我们预计本文描述的重组病毒将有助于研究广泛的 CD4 T 细胞反应。