Center for Eukaryotic Gene Regulation, Department of Biochemistry and Molecular Biology, The Pennsylvania State University, University Park, PA, USA.
Department of Molecular Biology and Genetics, Cornell University, Ithaca, NY, USA.
Nature. 2021 Apr;592(7853):309-314. doi: 10.1038/s41586-021-03314-8. Epub 2021 Mar 10.
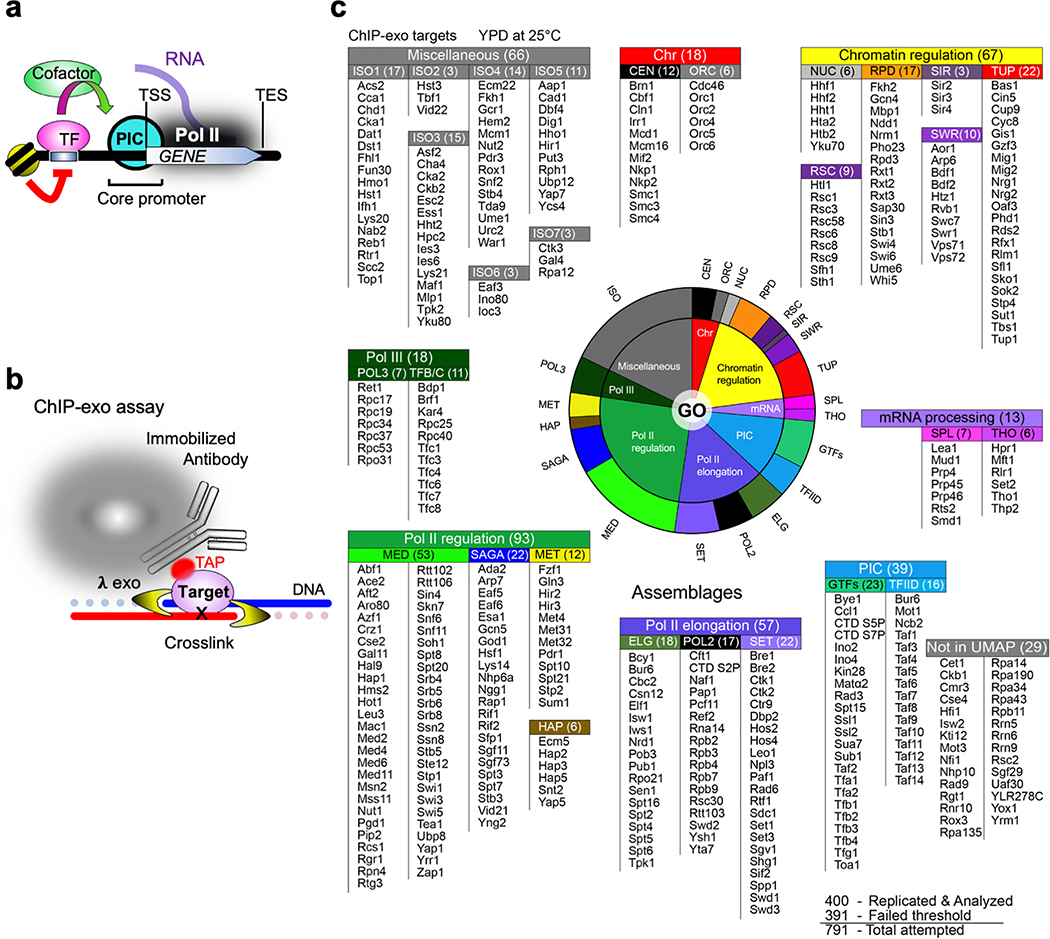
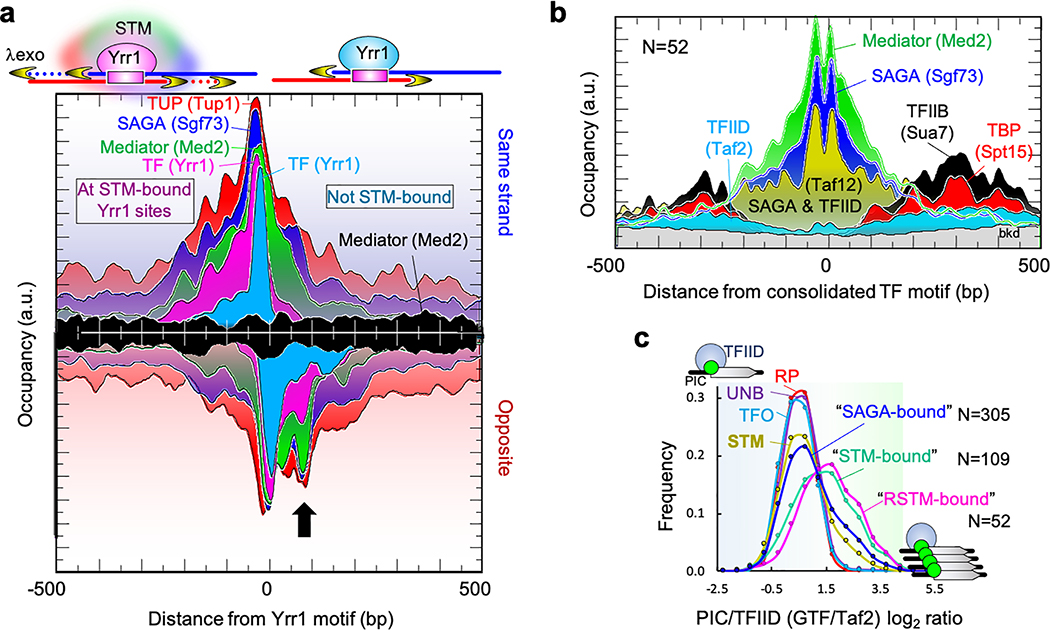
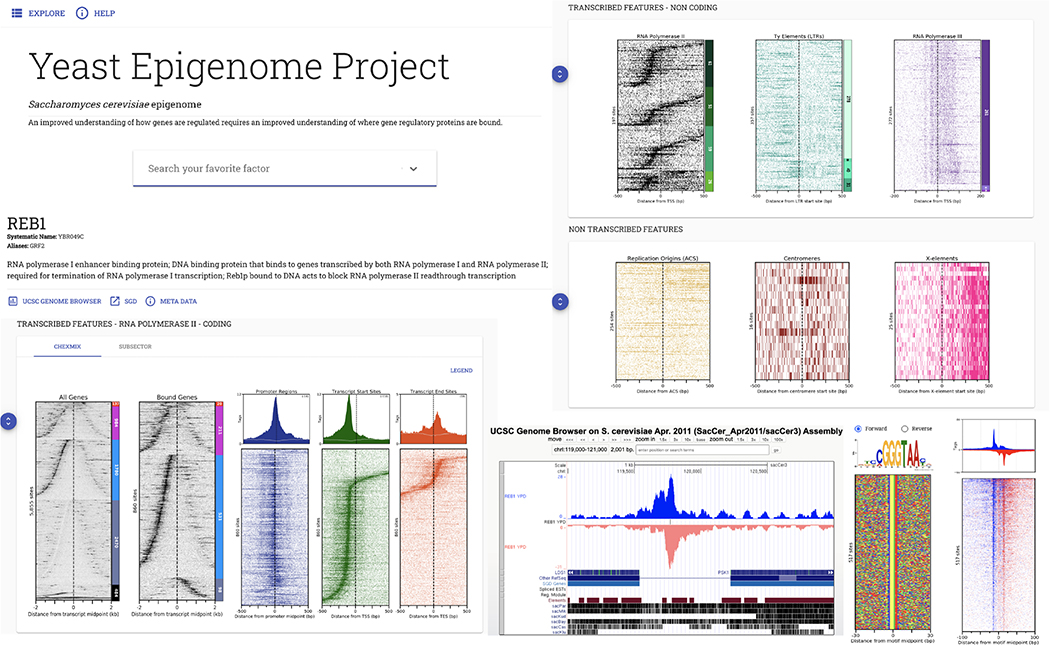
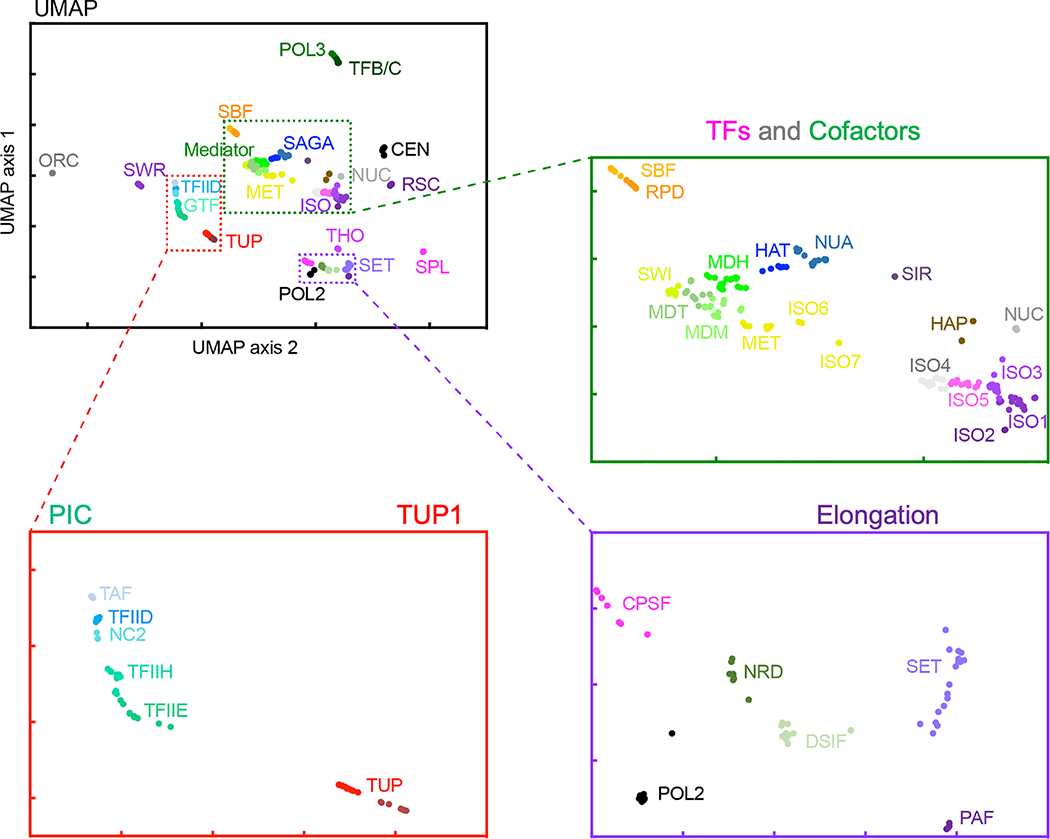
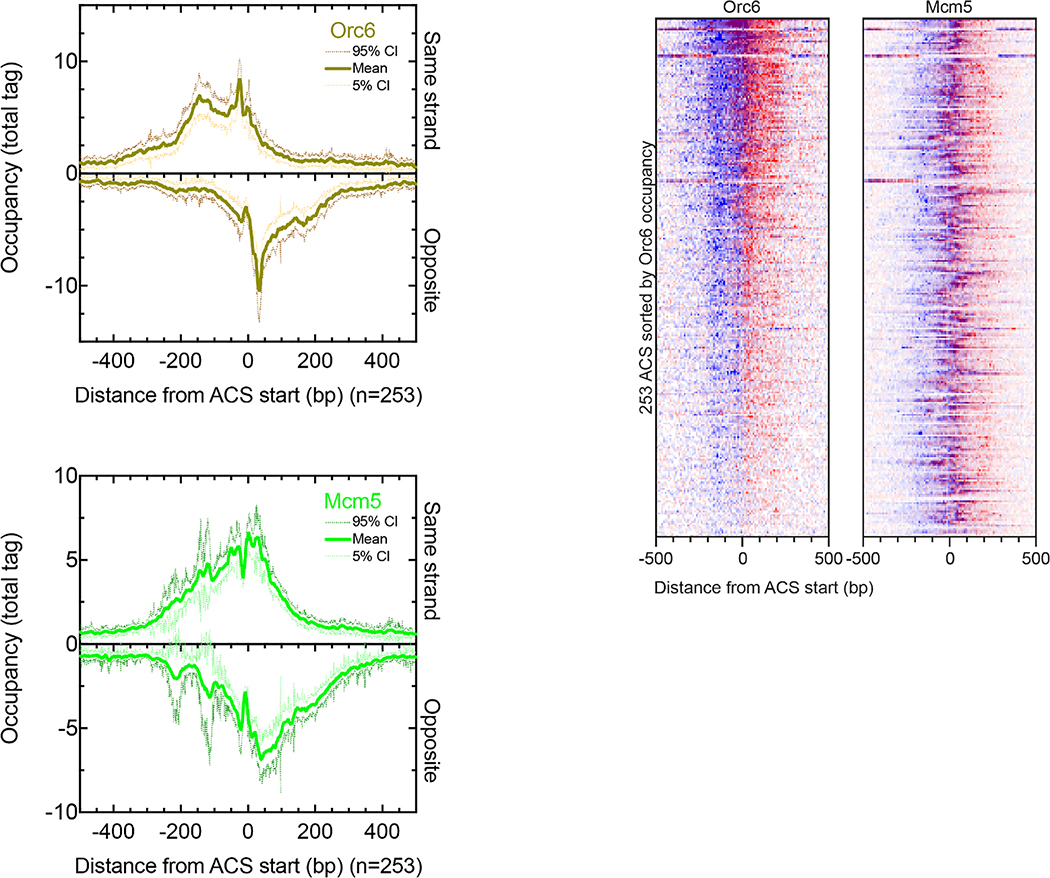
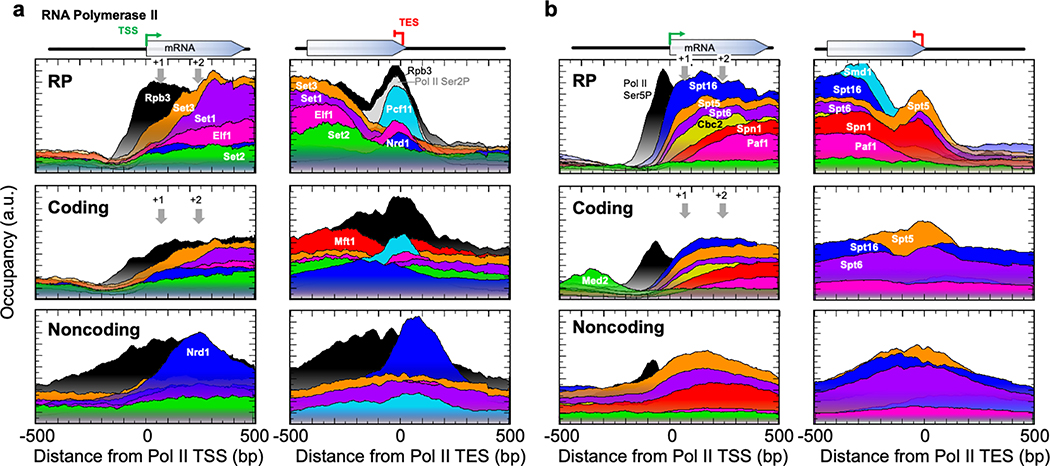
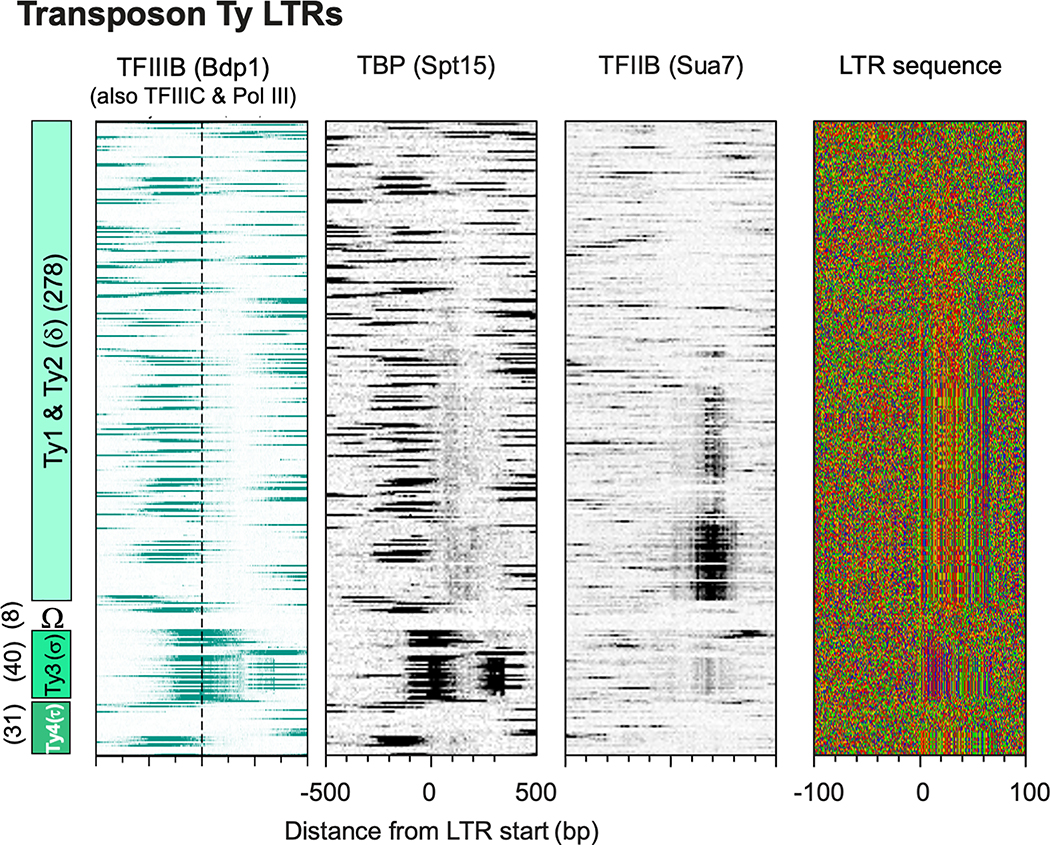
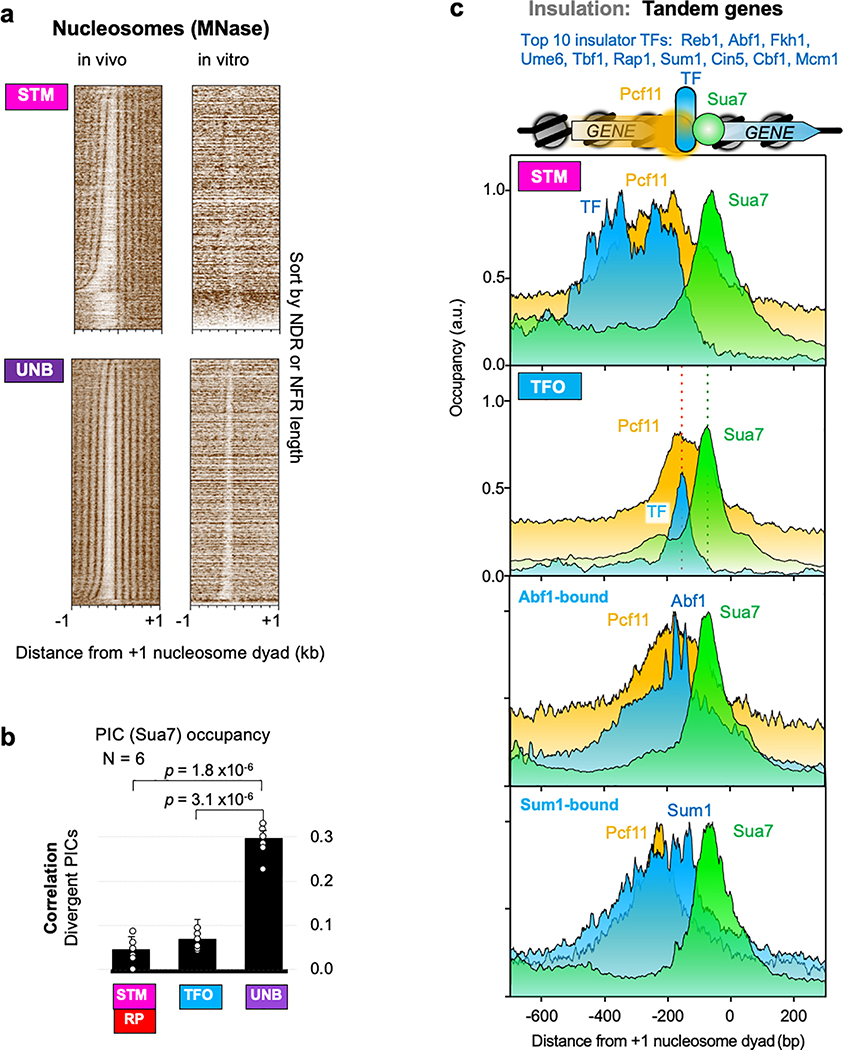
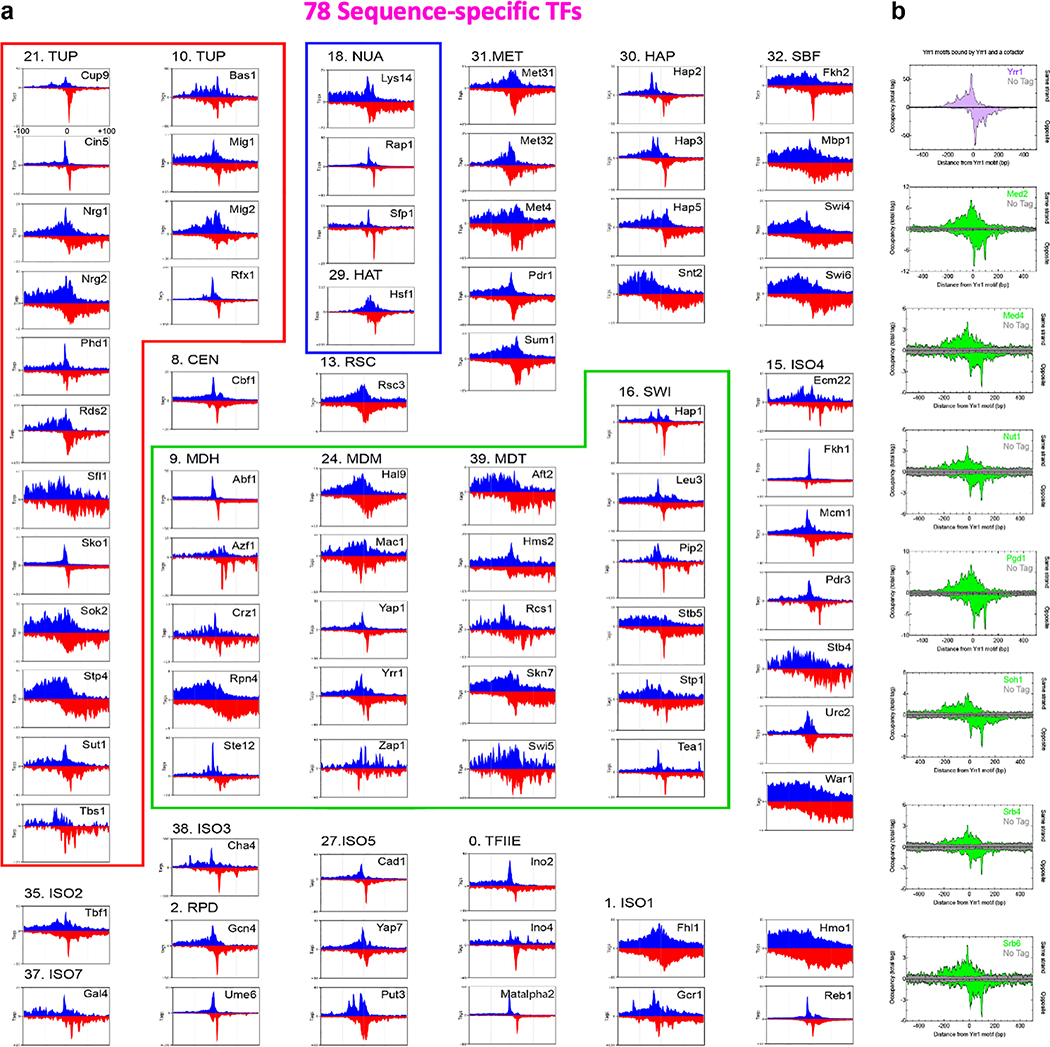
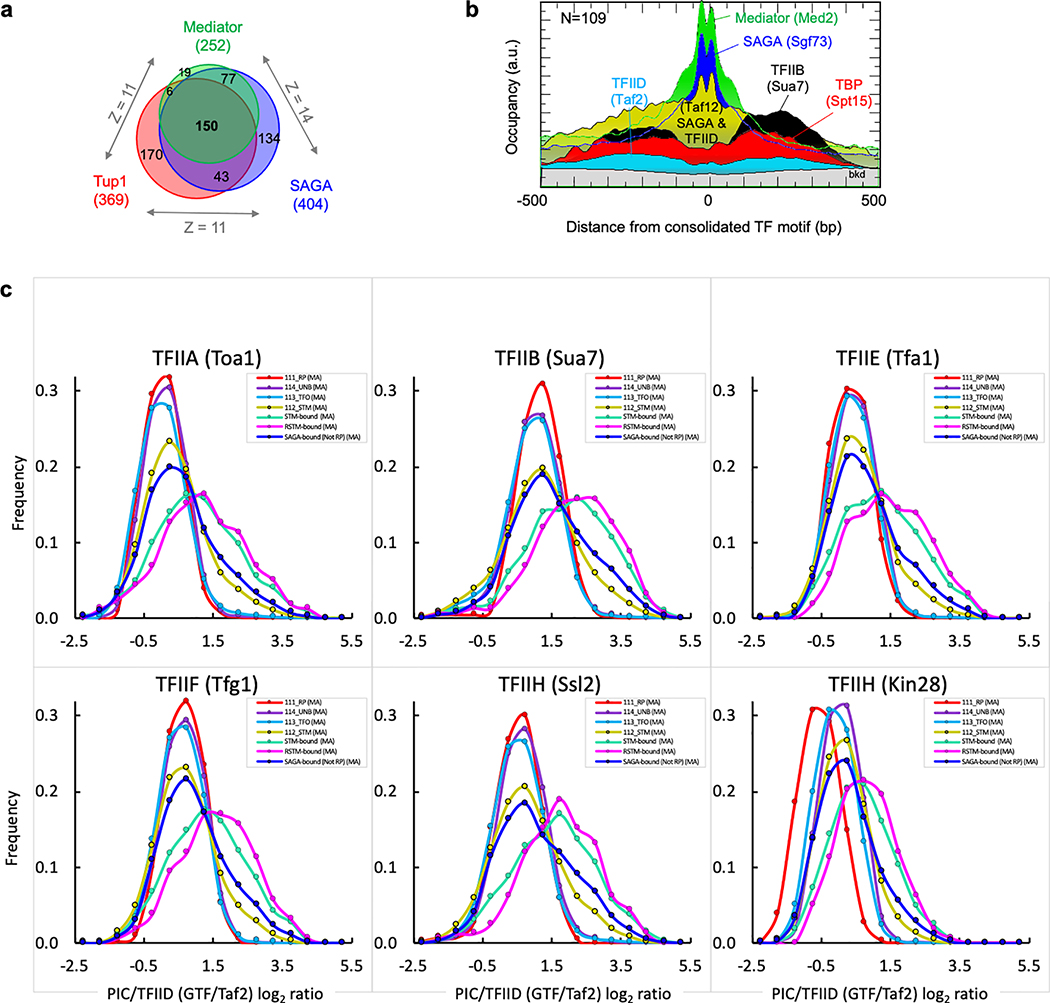
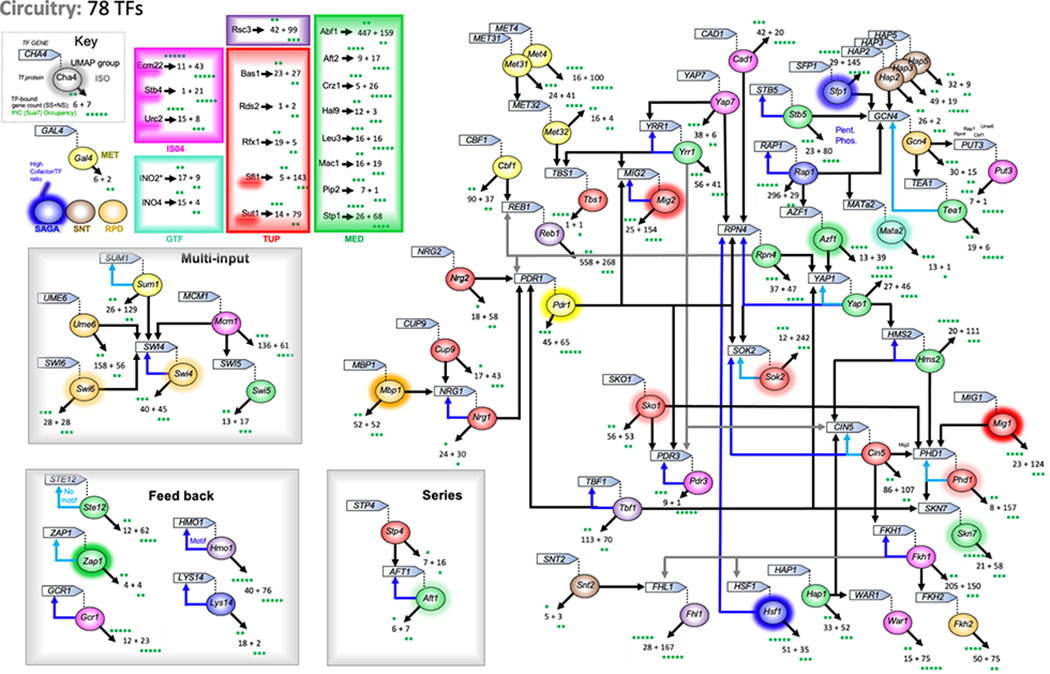
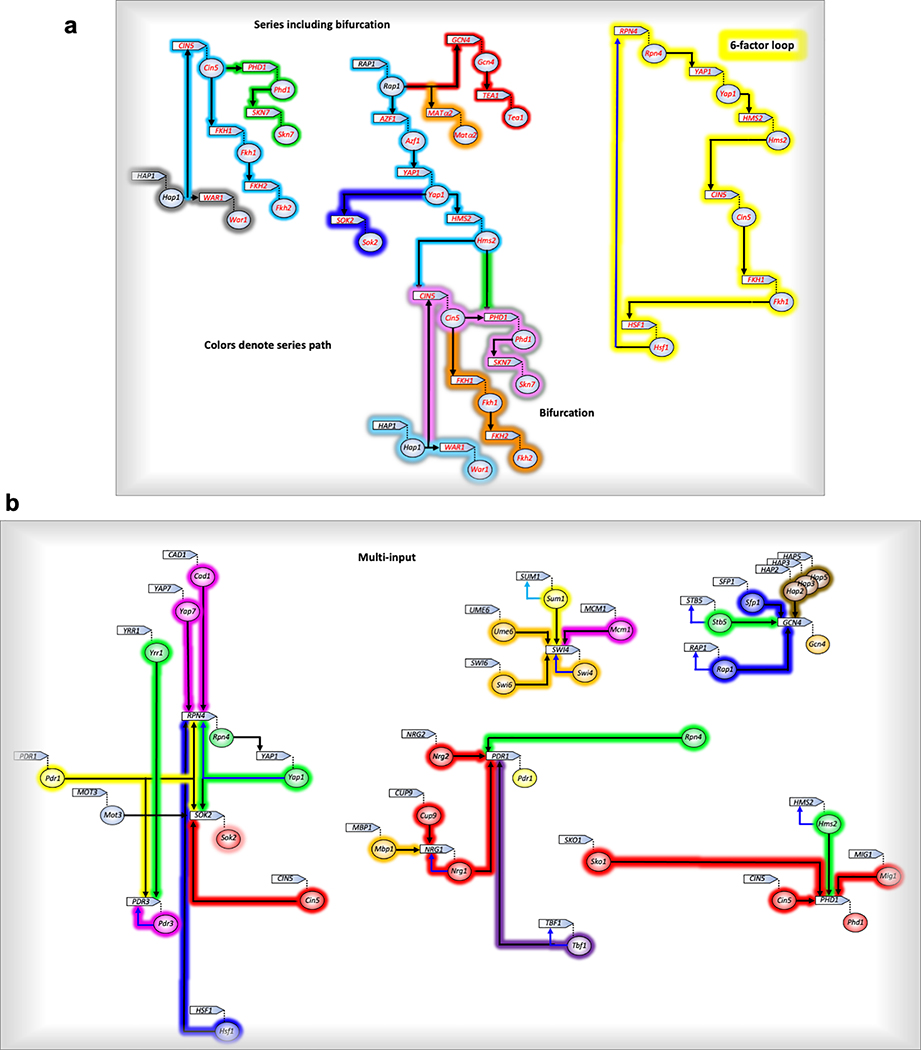
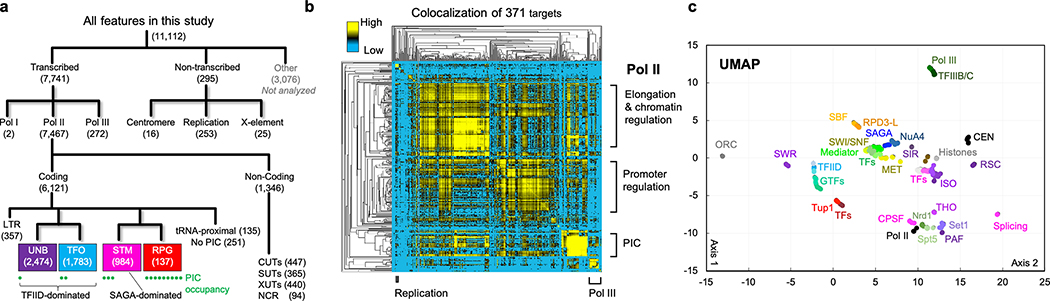
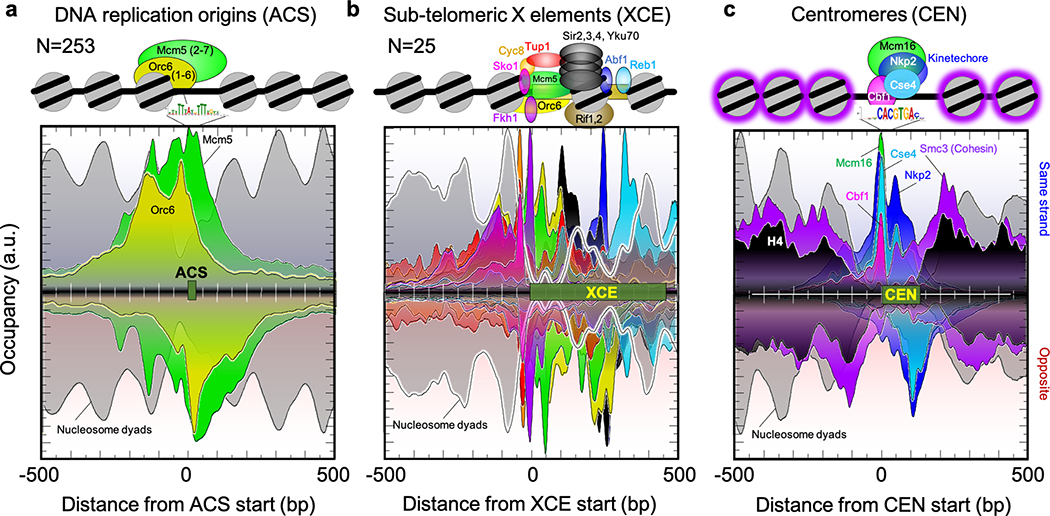
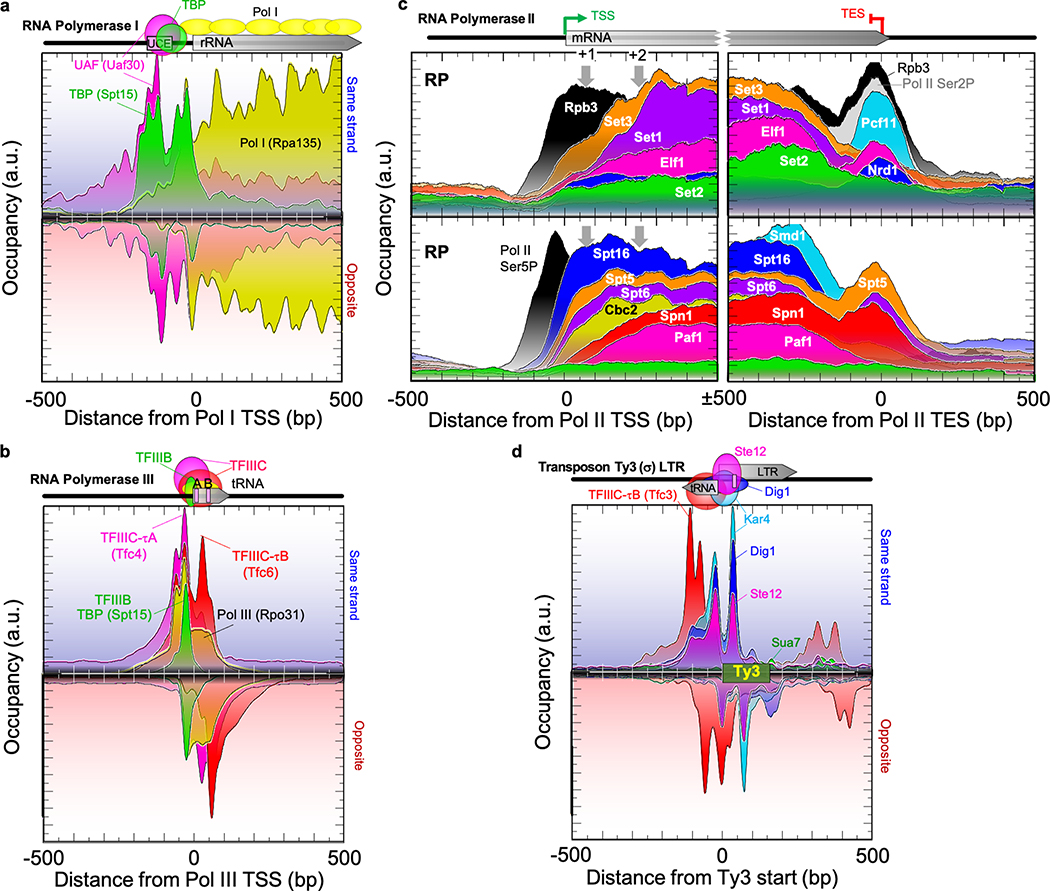
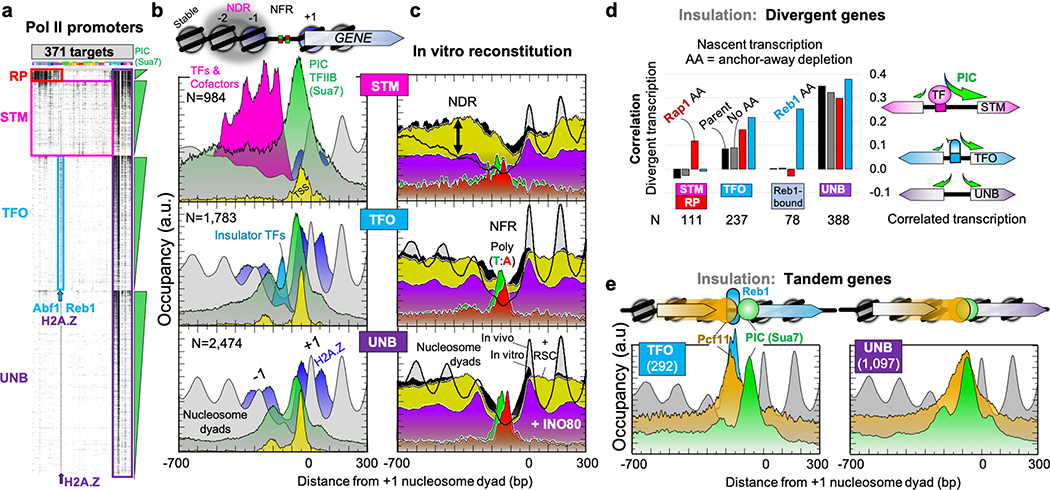
The genome-wide architecture of chromatin-associated proteins that maintains chromosome integrity and gene regulation is not well defined. Here we use chromatin immunoprecipitation, exonuclease digestion and DNA sequencing (ChIP-exo/seq) to define this architecture in Saccharomyces cerevisiae. We identify 21 meta-assemblages consisting of roughly 400 different proteins that are related to DNA replication, centromeres, subtelomeres, transposons and transcription by RNA polymerase (Pol) I, II and III. Replication proteins engulf a nucleosome, centromeres lack a nucleosome, and repressive proteins encompass three nucleosomes at subtelomeric X-elements. We find that most promoters associated with Pol II evolved to lack a regulatory region, having only a core promoter. These constitutive promoters comprise a short nucleosome-free region (NFR) adjacent to a +1 nucleosome, which together bind the transcription-initiation factor TFIID to form a preinitiation complex. Positioned insulators protect core promoters from upstream events. A small fraction of promoters evolved an architecture for inducibility, whereby sequence-specific transcription factors (ssTFs) create a nucleosome-depleted region (NDR) that is distinct from an NFR. We describe structural interactions among ssTFs, their cognate cofactors and the genome. These interactions include the nucleosomal and transcriptional regulators RPD3-L, SAGA, NuA4, Tup1, Mediator and SWI-SNF. Surprisingly, we do not detect interactions between ssTFs and TFIID, suggesting that such interactions do not stably occur. Our model for gene induction involves ssTFs, cofactors and general factors such as TBP and TFIIB, but not TFIID. By contrast, constitutive transcription involves TFIID but not ssTFs engaged with their cofactors. From this, we define a highly integrated network of gene regulation by ssTFs.
染色质相关蛋白的全基因组结构对于维持染色体完整性和基因调控至关重要,但目前其具体结构仍未完全明确。在这里,我们使用染色质免疫沉淀、核酸外切酶消化和 DNA 测序(ChIP-exo/seq)的方法,在酿酒酵母中定义了这种结构。我们确定了 21 个元组装体,其中包含大约 400 种不同的蛋白质,这些蛋白质与 DNA 复制、着丝粒、端粒、转座子和 RNA 聚合酶(Pol)I、II 和 III 转录有关。复制蛋白包裹核小体,着丝粒不含核小体,而抑制蛋白则围绕端粒 X 元件上的三个核小体。我们发现,大多数与 Pol II 相关的启动子已经进化到缺乏调节区,只有核心启动子。这些组成型启动子包含一个紧邻+1 核小体的短无核小体区(NFR),该区域与转录起始因子 TFIID 结合形成起始前复合物。定位的绝缘子保护核心启动子免受上游事件的影响。一小部分启动子进化出了可诱导的结构,其中序列特异性转录因子(ssTF)创建一个与 NFR 不同的无核小体区(NDR)。我们描述了 ssTF、其同源辅助因子和基因组之间的结构相互作用。这些相互作用包括核小体和转录调节因子 RPD3-L、SAGA、NuA4、Tup1、共激活因子和 SWI-SNF。令人惊讶的是,我们没有检测到 ssTF 与 TFIID 之间的相互作用,这表明这种相互作用不会稳定发生。我们的基因诱导模型涉及 ssTF、辅助因子和通用因子,如 TBP 和 TFIIB,但不涉及 TFIID。相比之下,组成型转录涉及 TFIID,但不涉及与辅助因子结合的 ssTF。由此,我们定义了一个由 ssTF 介导的高度综合的基因调控网络。