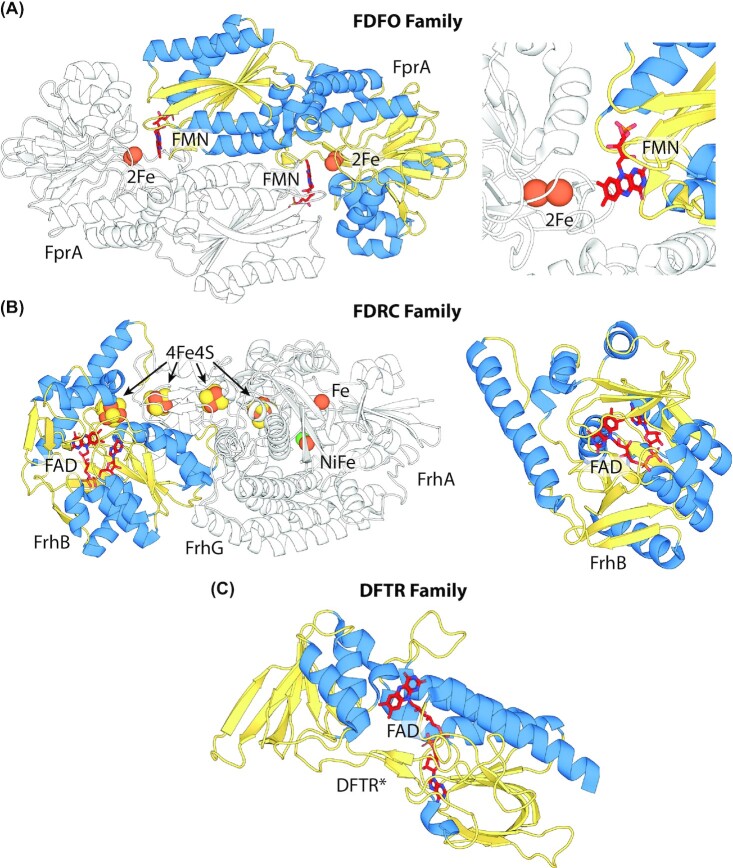
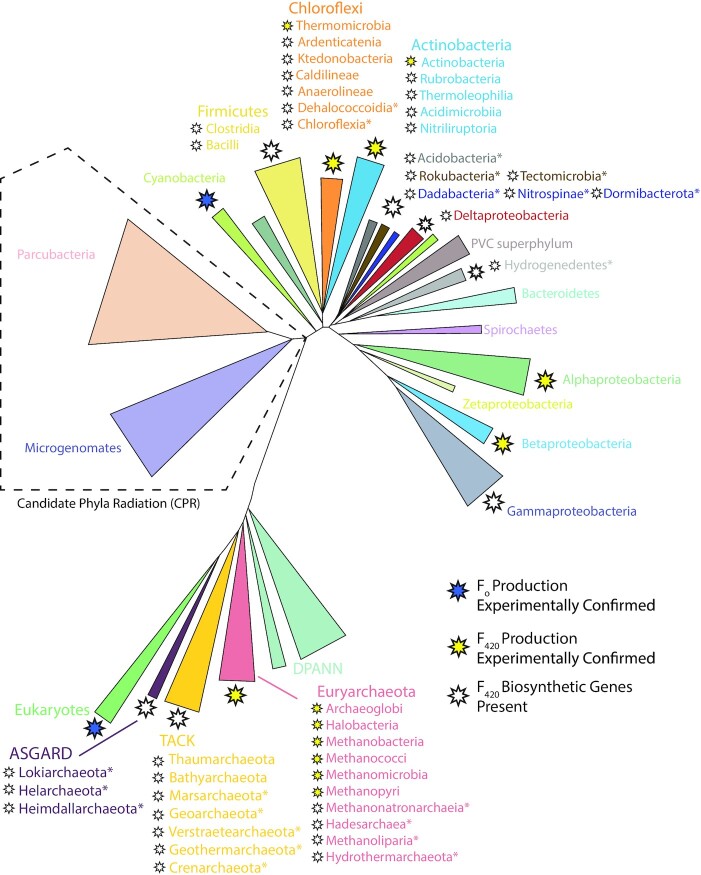
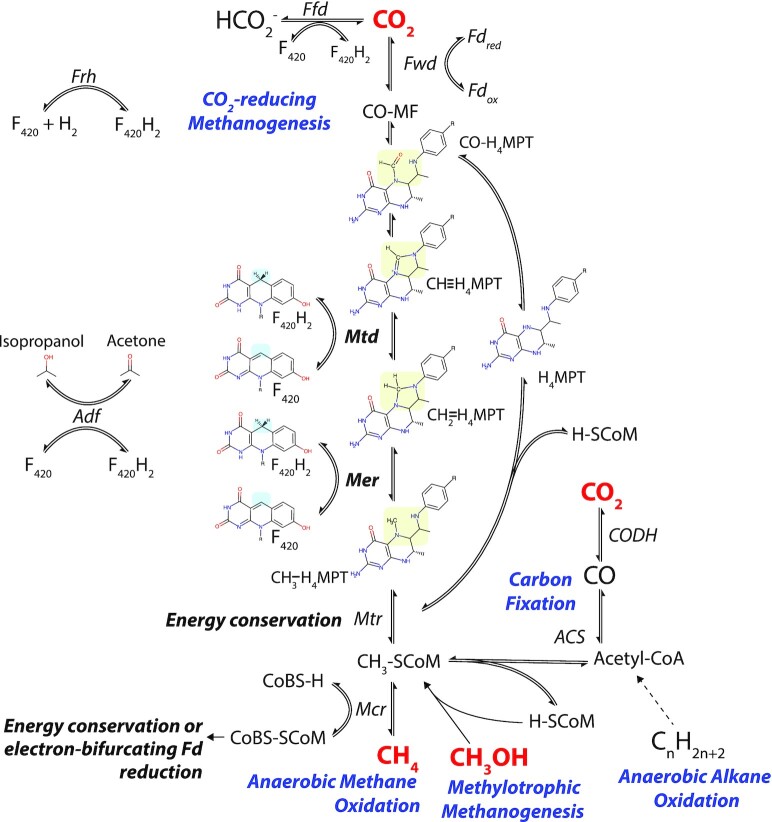
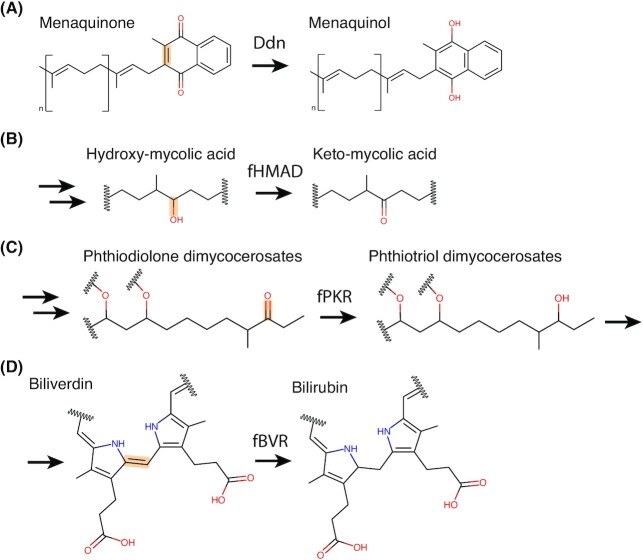
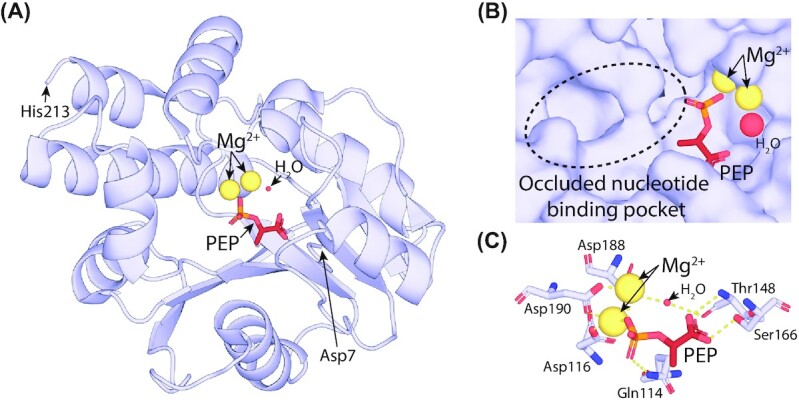
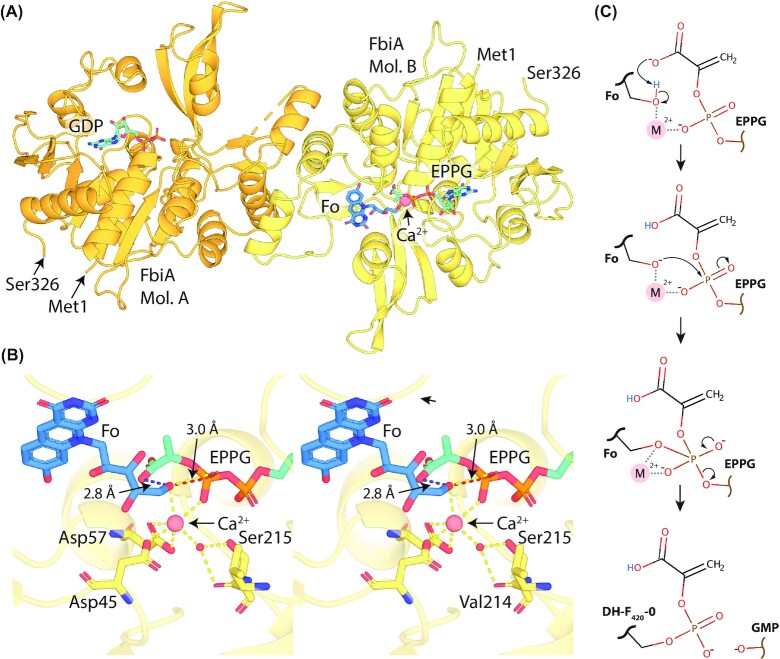
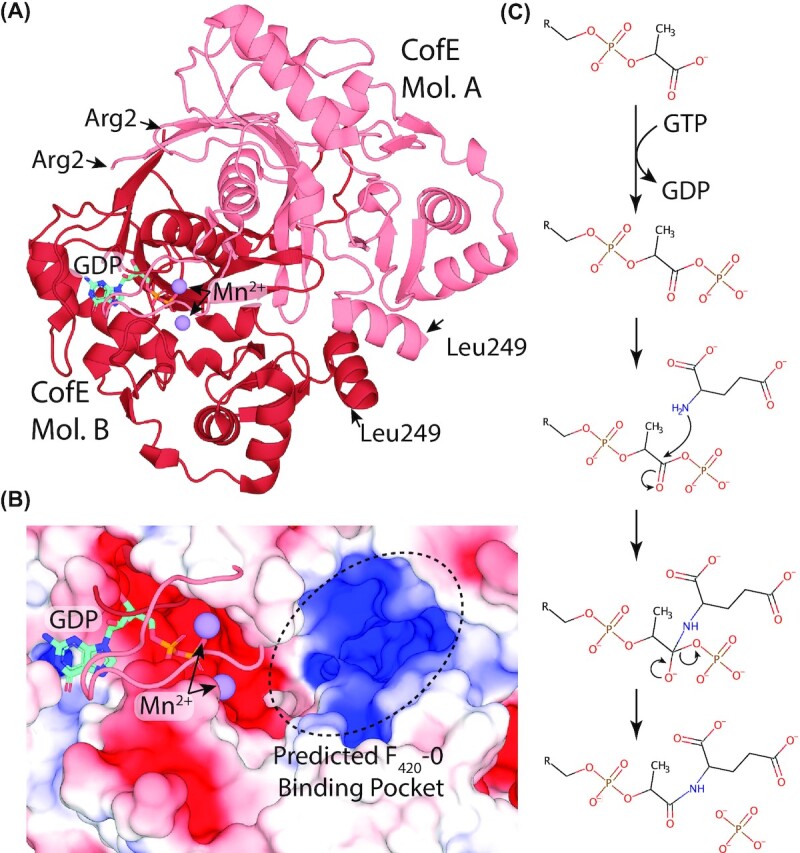
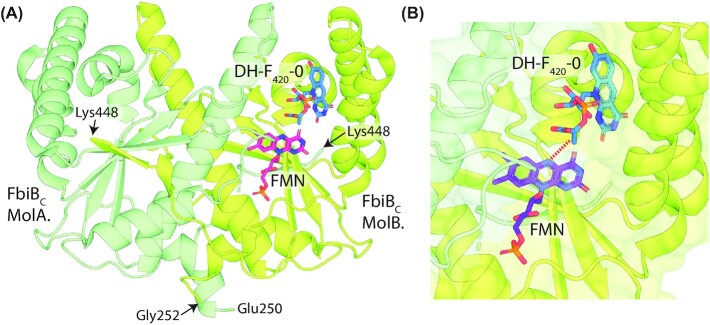
Department of Microbiology, Monash Biomedicine Discovery Institute, Monash University, Clayton, VIC 3800, Australia.
FEMS Microbiol Rev. 2021 Sep 8;45(5). doi: 10.1093/femsre/fuab021.
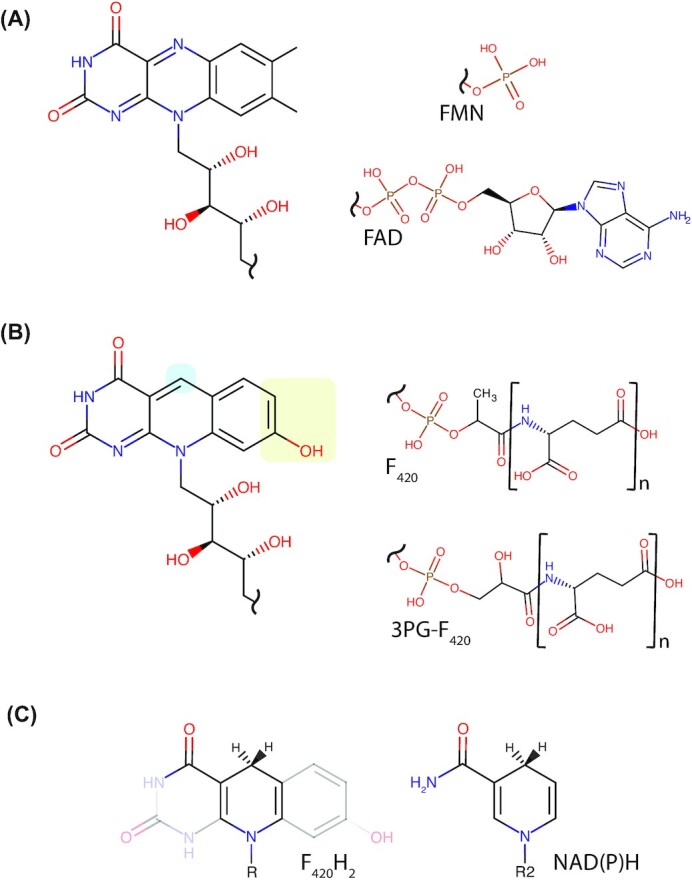
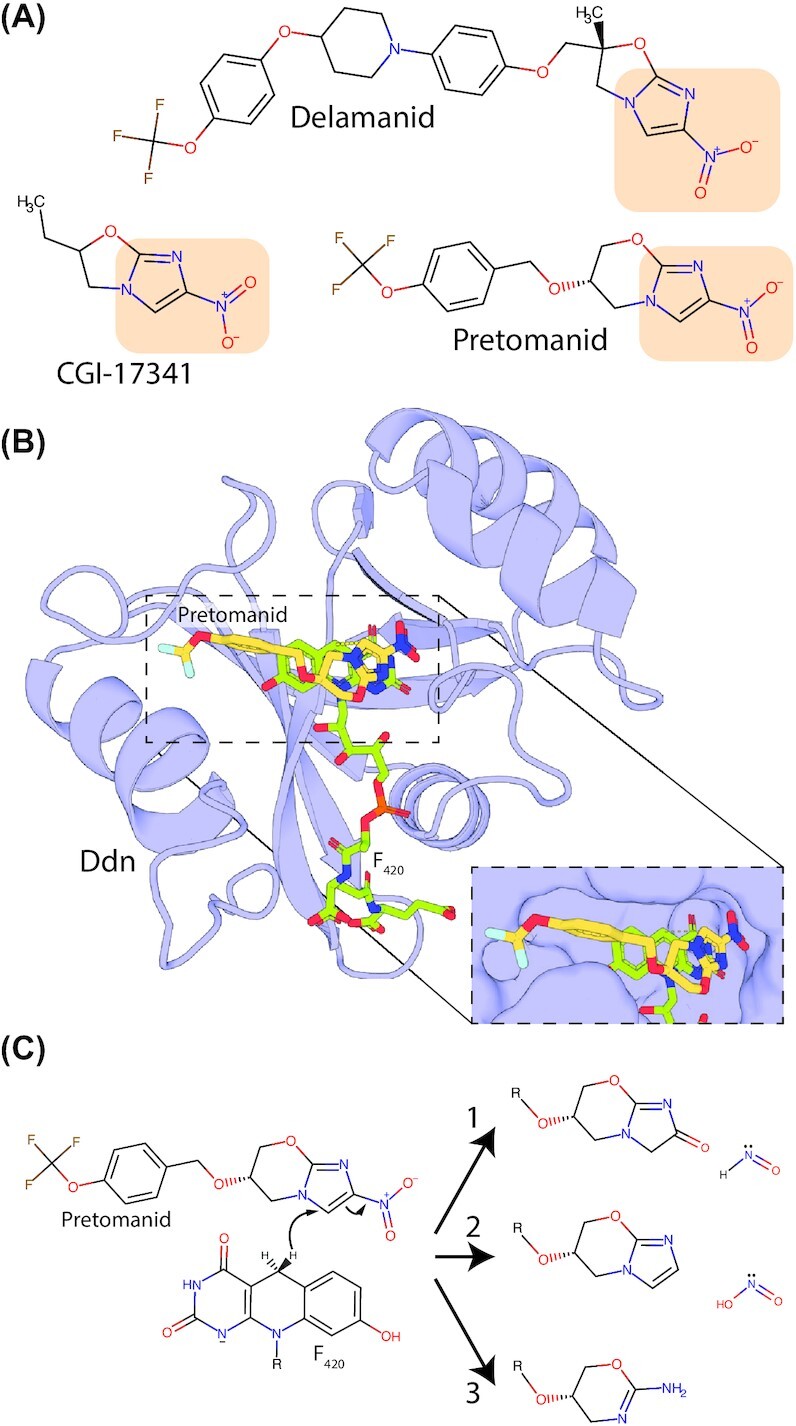
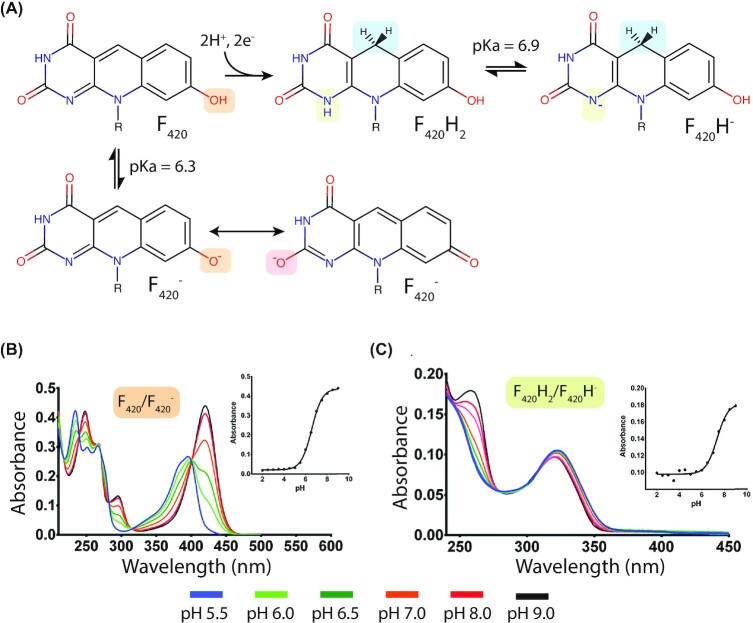
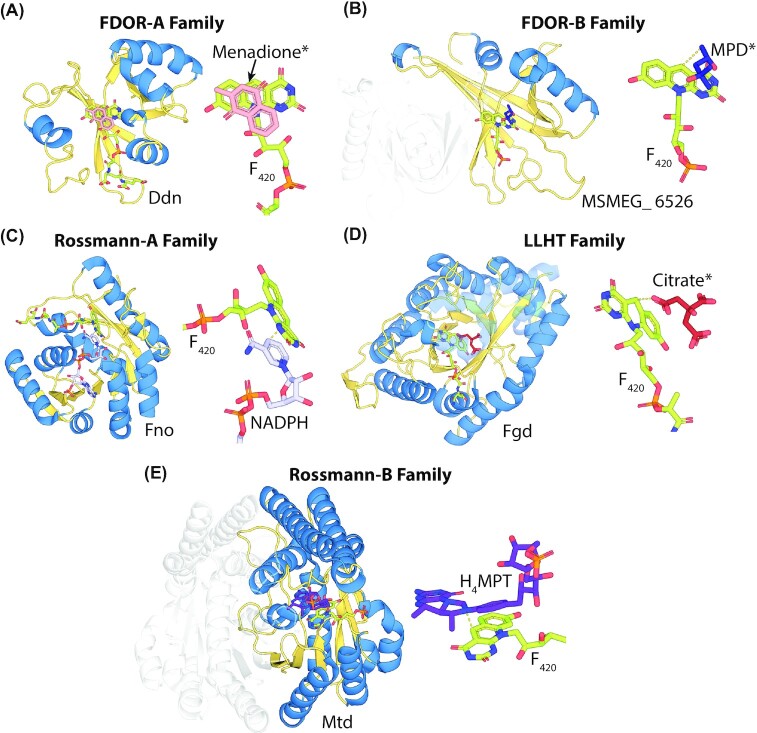
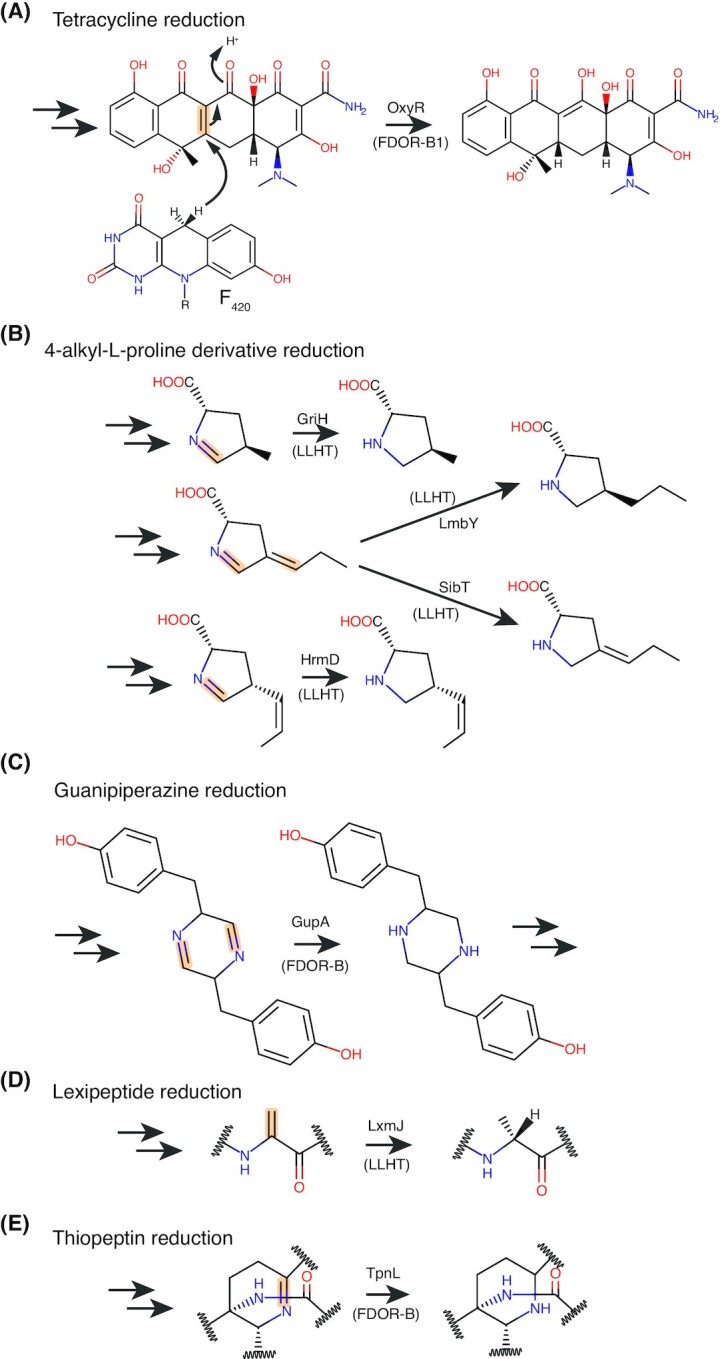
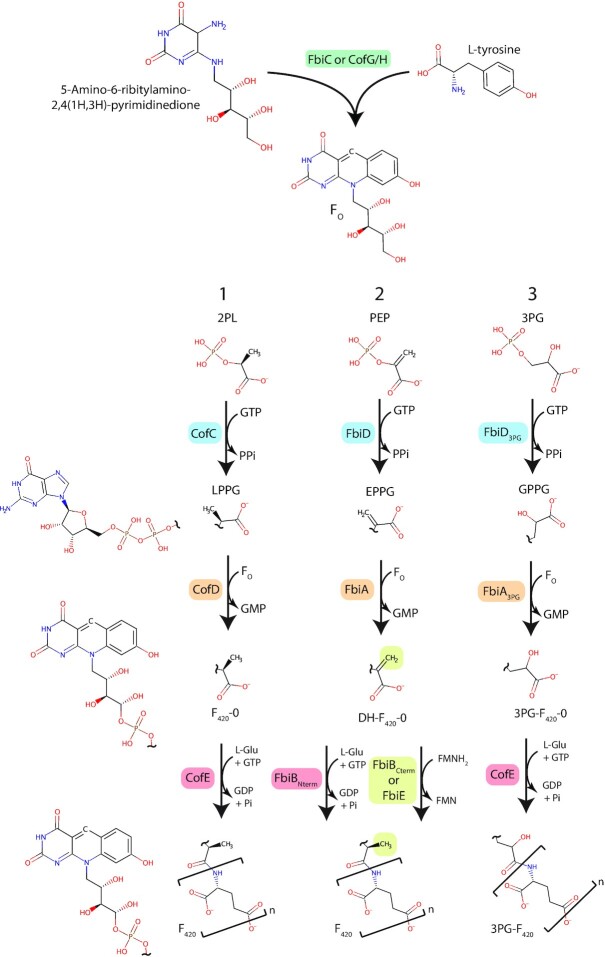
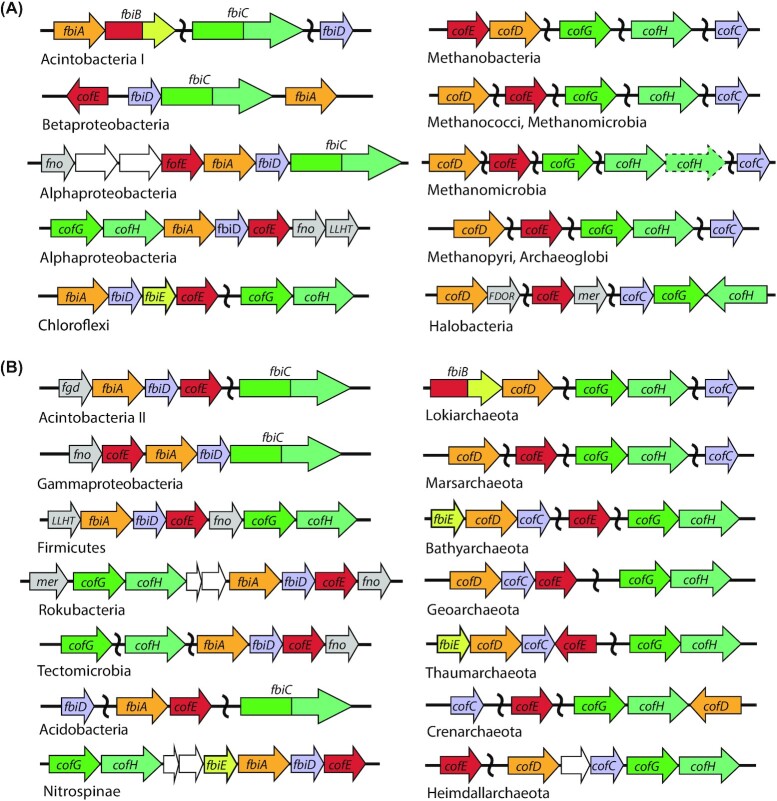
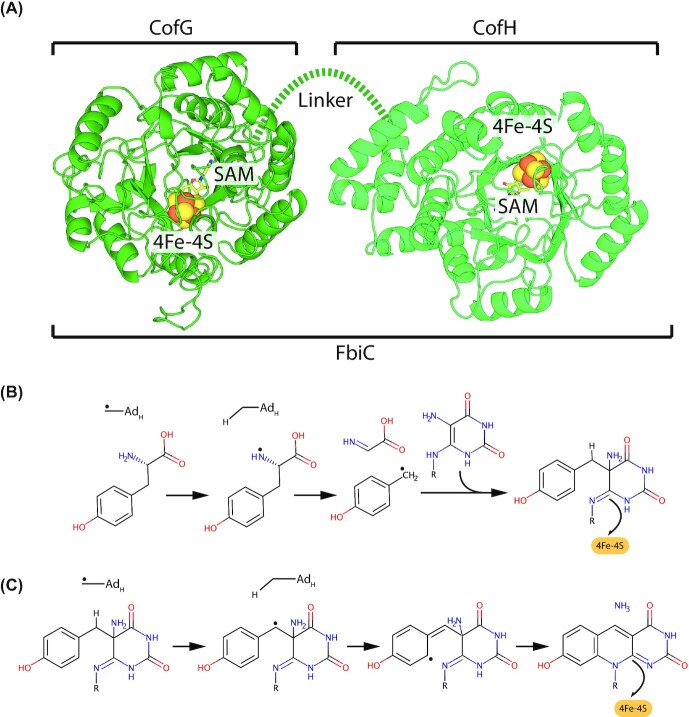
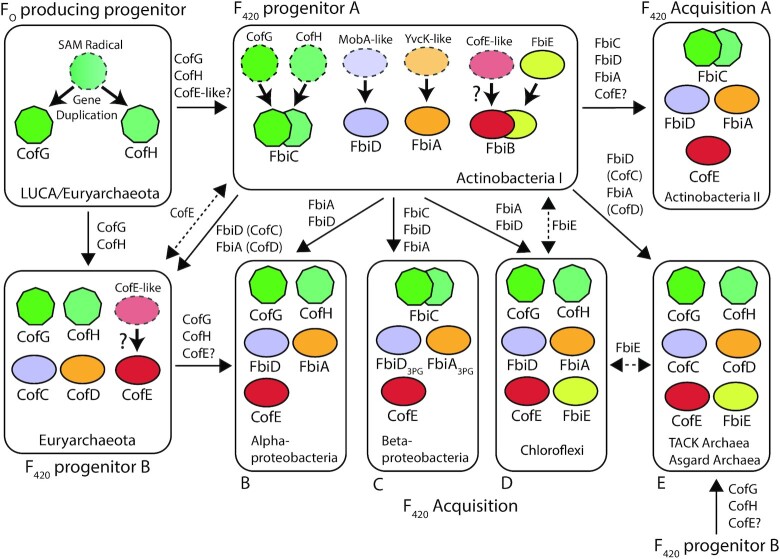
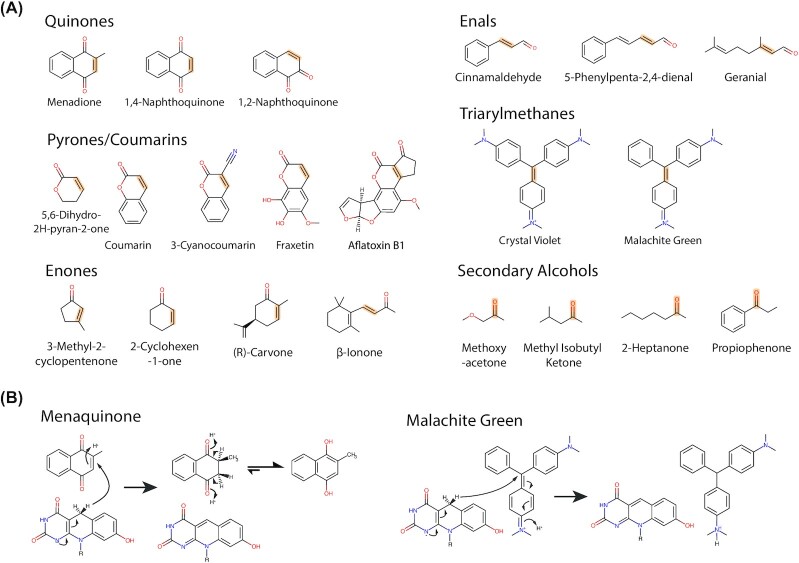
Many bacteria and archaea produce the redox cofactor F420. F420 is structurally similar to the cofactors FAD and FMN but is catalytically more similar to NAD and NADP. These properties allow F420 to catalyze challenging redox reactions, including key steps in methanogenesis, antibiotic biosynthesis and xenobiotic biodegradation. In the last 5 years, there has been much progress in understanding its distribution, biosynthesis, role and applications. Whereas F420 was previously thought to be confined to Actinobacteria and Euryarchaeota, new evidence indicates it is synthesized across the bacterial and archaeal domains, as a result of extensive horizontal and vertical biosynthetic gene transfer. F420 was thought to be synthesized through one biosynthetic pathway; however, recent advances have revealed variants of this pathway and have resolved their key biosynthetic steps. In parallel, new F420-dependent biosynthetic and metabolic processes have been discovered. These advances have enabled the heterologous production of F420 and identified enantioselective F420H2-dependent reductases for biocatalysis. New research has also helped resolve how microorganisms use F420 to influence human and environmental health, providing opportunities for tuberculosis treatment and methane mitigation. A total of 50 years since its discovery, multiple paradigms associated with F420 have shifted, and new F420-dependent organisms and processes continue to be discovered.
许多细菌和古菌都能产生氧化还原辅因子 F420。F420 在结构上与辅因子 FAD 和 FMN 相似,但在催化上更类似于 NAD 和 NADP。这些特性使 F420 能够催化具有挑战性的氧化还原反应,包括甲烷生成、抗生素生物合成和异生物质生物降解中的关键步骤。在过去的 5 年中,人们在理解其分布、生物合成、作用和应用方面取得了很大进展。虽然 F420 以前被认为仅限于放线菌和广古菌,但新的证据表明,由于广泛的水平和垂直生物合成基因转移,它在细菌和古菌领域中都有合成。F420 被认为是通过一条生物合成途径合成的;然而,最近的进展揭示了这条途径的变体,并解决了其关键的生物合成步骤。与此同时,新的依赖 F420 的生物合成和代谢过程也被发现。这些进展使 F420 的异源生产成为可能,并鉴定了用于生物催化的对映选择性 F420H2 依赖性还原酶。新的研究还有助于解决微生物如何利用 F420 来影响人类和环境健康,为结核病治疗和甲烷减排提供了机会。自发现以来已经过去了 50 年,与 F420 相关的多个范式已经发生了转变,并且不断有新的依赖 F420 的生物体和过程被发现。