Institute of Immunology, Charité-Universitätsmedizin Berlin, corporate member of Freie Universität Berlin and Humboldt-Universität zu Berlin, Berlin, Germany.
German Cancer Research Center, Heidelberg, Germany.
Elife. 2021 Apr 20;10:e62019. doi: 10.7554/eLife.62019.
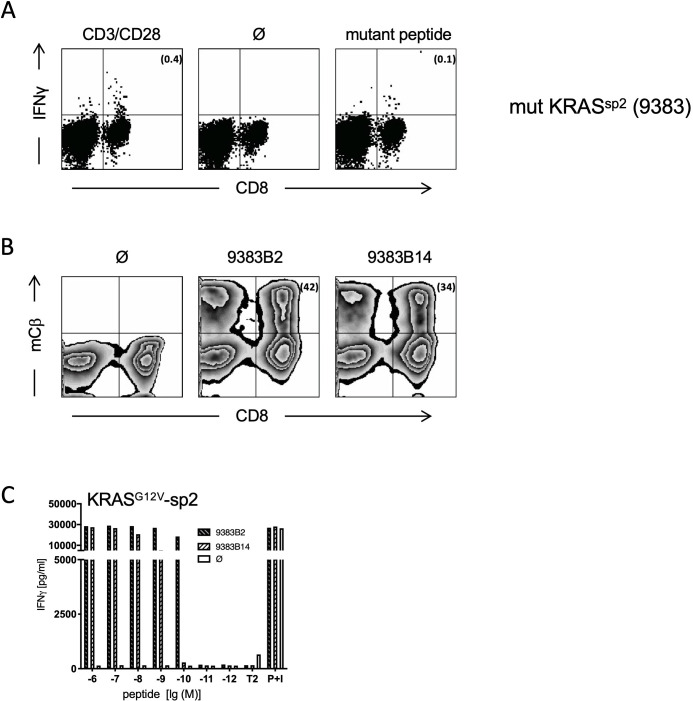
Proteasome-catalyzed peptide splicing (PCPS) of cancer-driving antigens could generate attractive neoepitopes to be targeted by T cell receptor (TCR)-based adoptive T cell therapy. Based on a spliced peptide prediction algorithm, TCRs were generated against putative KRAS- and RAC2-derived neo-splicetopes with high HLA-A*02:01 binding affinity. TCRs generated in mice with a diverse human TCR repertoire specifically recognized the respective target peptides with high efficacy. However, we failed to detect any neo-splicetope-specific T cell response when testing the in vivo neo-splicetope generation and obtained no experimental evidence that the putative KRAS- and RAC2-derived neo-splicetopes were naturally processed and presented. Furthermore, only the putative RAC2-derived neo-splicetopes was generated by in vitro PCPS. The experiments pose severe questions on the notion that available algorithms or the in vitro PCPS reaction reliably simulate in vivo splicing and argue against the general applicability of an algorithm-driven 'reverse immunology' pipeline for the identification of cancer-specific neo-splicetopes.
蛋白酶体催化的癌驱动抗原肽剪接(PCPS)可以产生有吸引力的新表位,以作为 TCR 为基础的过继性 T 细胞治疗的靶点。基于一个剪接肽预测算法,我们针对具有高 HLA-A*02:01 结合亲和力的潜在 KRAS 和 RAC2 衍生的新剪接表位生成了 TCR。在具有多样化人类 TCR repertoire 的小鼠中生成的 TCR 特异性地以高功效识别各自的靶肽。然而,当测试体内新剪接表位的产生时,我们未能检测到任何新剪接表位特异性 T 细胞反应,并且没有获得实验证据表明潜在的 KRAS 和 RAC2 衍生的新剪接表位是天然加工和呈递的。此外,只有潜在的 RAC2 衍生的新剪接表位是通过体外 PCPS 产生的。这些实验对以下观点提出了严峻的质疑,即现有的算法或体外 PCPS 反应是否能可靠地模拟体内剪接,并反对基于算法的“反向免疫学法”管道用于鉴定癌症特异性新剪接表位的一般适用性。