Division of Biological Sciences, University of Montana, Missoula Montana, United States of America.
Keck Science Department, Claremont-McKenna, Scripps, and Pitzer Colleges, Claremont California, United States of America.
PLoS Genet. 2021 Apr 22;17(4):e1009418. doi: 10.1371/journal.pgen.1009418. eCollection 2021 Apr.
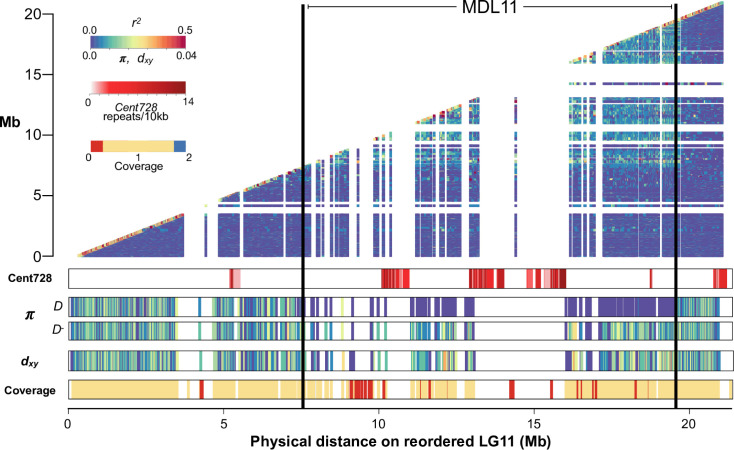
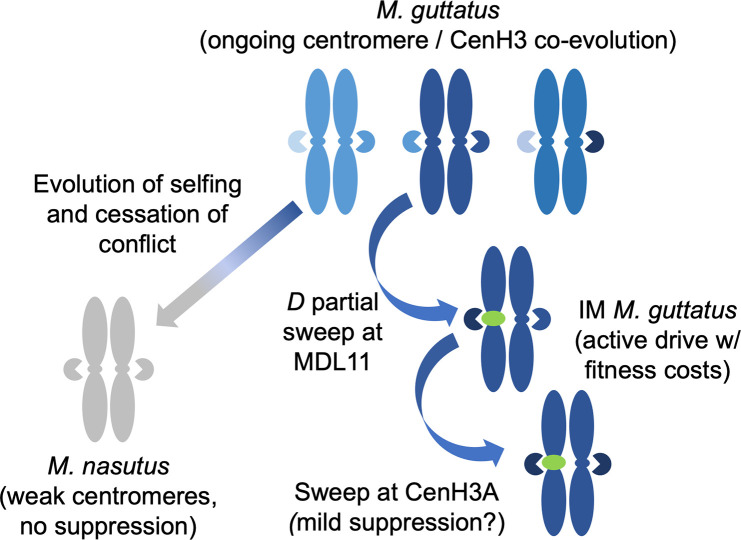
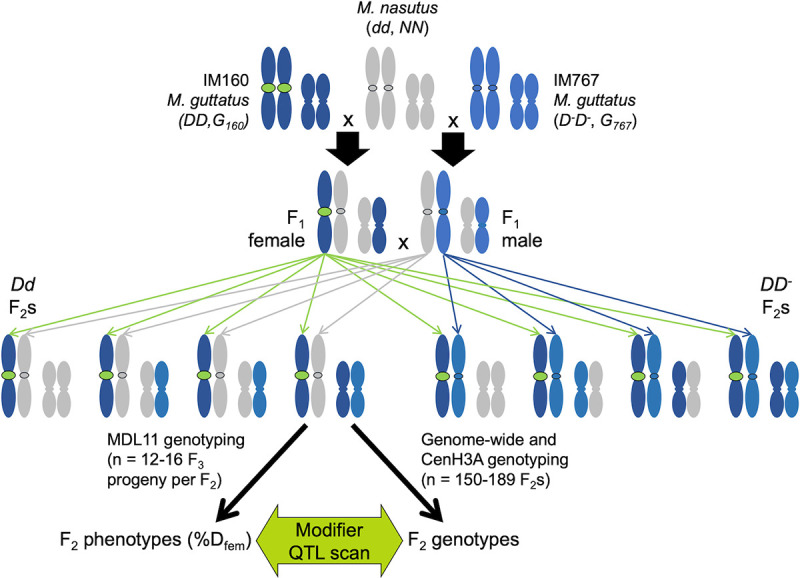
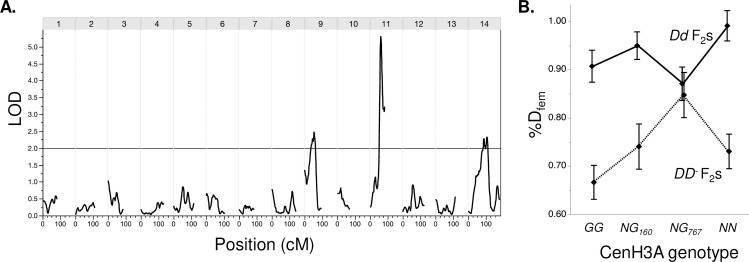
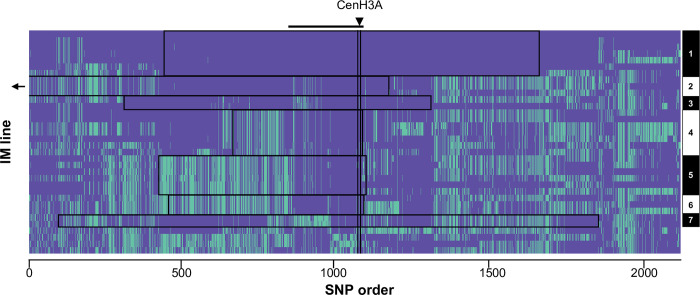
Centromeres are essential mediators of chromosomal segregation, but both centromeric DNA sequences and associated kinetochore proteins are paradoxically diverse across species. The selfish centromere model explains rapid evolution by both components via an arms-race scenario: centromeric DNA variants drive by distorting chromosomal transmission in female meiosis and attendant fitness costs select on interacting proteins to restore Mendelian inheritance. Although it is clear than centromeres can drive and that drive often carries costs, female meiotic drive has not been directly linked to selection on kinetochore proteins in any natural system. Here, we test the selfish model of centromere evolution in a yellow monkeyflower (Mimulus guttatus) population polymorphic for a costly driving centromere (D). We show that the D haplotype is structurally and genetically distinct and swept to a high stable frequency within the past 1500 years. We use quantitative genetic mapping to demonstrate that context-dependence in the strength of drive (from near-100% D transmission in interspecific hybrids to near-Mendelian in within-population crosses) primarily reflects variable vulnerability of the non-driving competitor chromosomes, but also map an unlinked modifier of drive coincident with kinetochore protein Centromere-specific Histone 3 A (CenH3A). Finally, CenH3A exhibits a recent (<1000 years) selective sweep in our focal population, implicating local interactions with D in ongoing adaptive evolution of this kinetochore protein. Together, our results demonstrate an active co-evolutionary arms race between DNA and protein components of the meiotic machinery in Mimulus, with important consequences for individual fitness and molecular divergence.
着丝粒是染色体分离的重要介质,但着丝粒 DNA 序列和相关的动粒蛋白在物种间都是多种多样的,这是一个悖论。自私的着丝粒模型通过军备竞赛场景解释了这两个组成部分的快速进化:着丝粒 DNA 变体通过扭曲雌性减数分裂中的染色体传递,并伴随着适合度成本,选择相互作用的蛋白质来恢复孟德尔遗传。尽管很明显,着丝粒可以驱动,并且驱动通常伴随着成本,但在任何自然系统中,雌性减数分裂驱动都没有直接与动粒蛋白的选择联系起来。在这里,我们在一个黄色的风铃草(Mimulus guttatus)种群中测试了着丝粒进化的自私模型,该种群存在一个代价高昂的驱动着丝粒(D)。我们表明,D 单倍型在结构和遗传上是不同的,并在过去 1500 年内迅速达到了一个高的稳定频率。我们利用定量遗传作图来证明驱动强度的上下文依赖性(从种间杂种中近 100%的 D 传递到种群内杂交中的近孟德尔传递)主要反映了非驱动竞争染色体的易损性不同,但也映射了一个与动粒蛋白着丝粒特异性组蛋白 3A(CenH3A)不相关的驱动修饰因子。最后,CenH3A 在我们的焦点种群中表现出最近(<1000 年)的选择清除,这表明与 D 的局部相互作用在这个动粒蛋白的持续适应性进化中。总之,我们的结果表明,在风铃草中,减数分裂机制的 DNA 和蛋白质成分之间存在着积极的共同进化军备竞赛,这对个体适合度和分子分化有重要影响。