Department of Biology and Center for Biodiversity and Ecosystem Stewardship, Villanova University, Villanova, PA, USA.
Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI, USA.
BMC Ecol Evol. 2021 May 16;21(1):86. doi: 10.1186/s12862-021-01821-w.
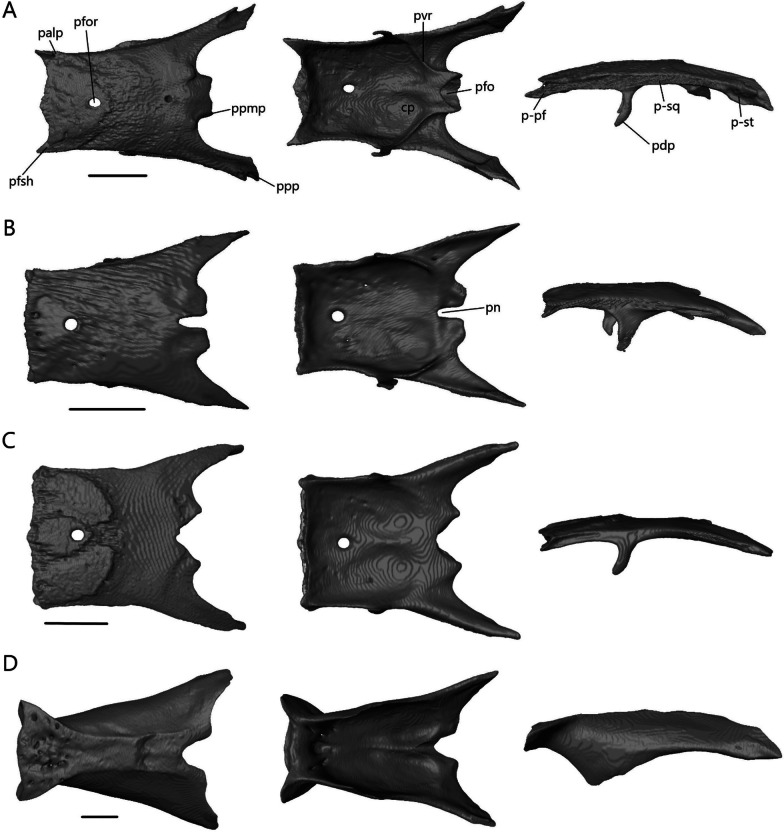
Skulls serve many functions and as a result, are subject to many different evolutionary pressures. In squamates, many fossorial species occupy a unique region of skull morphospace, showing convergence across families, due to modifications related to head-first burrowing. As different substrates have variable physical properties, particular skull shapes may offer selective advantages in certain substrates. Despite this, studies of variation within burrowers have been limited and are typically focused on a single origin of fossoriality. We focused on seven skink genera (Acontias, Typhlosaurus, Scelotes, Sepsina, Feylinia, Typhlacontias, and Mochlus; 39 sp.) from southern Africa, encompassing at least three independent evolutions of semi-fossoriality/fossoriality. We used microCT scans and geometric morphometrics to test how cranial and mandibular shape were influenced by phylogenetic history, size, and ecology. We also qualitatively described the skulls of four species to look at variation across phylogenetic and functional levels, and assess the degree of convergence.
We found a strong effect of phylogenetic history on cranial and mandibular shape, with size and substrate playing secondary roles. There was a clear gradient in morphospace from less specialized to more specialized burrowers and burrowers in sand were significantly different from those in other substrates. We also created an anatomical atlas for four species with each element described in isolation. Every bone showed some variation in shape and relative scaling of features, with the skull roofing bones, septomaxilla, vomer, and palatine showing the most variation. We showed how broad-scale convergence in traits related to fossoriality can be the result of different anatomical changes.
Our study used geometric morphometrics and comparative anatomy to examine how skull morphology changes for a highly specialized and demanding lifestyle. Although there was broad convergence in both shape and qualitative traits, phylogenetic history played a large role and much of this convergence was produced by different anatomical changes, implying different developmental pathways or lineage-specific constraints. Even within a single family, adaptation for a specialized ecology does not follow a singular deterministic path.
颅骨有多种功能,因此受到多种不同进化压力的影响。在有鳞目动物中,许多穴居物种占据颅骨形态空间的独特区域,由于与头部首先钻入有关的改造,表现出跨科的趋同。由于不同的基质具有不同的物理特性,特定的颅骨形状可能在某些基质中提供选择优势。尽管如此,对穴居动物内部变异的研究仍然有限,通常集中在单一的穴居起源上。我们专注于来自南非的七个石龙子属(Acontias、Typhlosaurus、Scelotes、Sepsina、Feylinia、Typhlacontias 和 Mochlus;39 种),包括至少三个半穴居/穴居的独立进化。我们使用微 CT 扫描和几何形态测量学来测试颅骨和下颌骨形状如何受到系统发育历史、体型和生态的影响。我们还定性描述了四个物种的颅骨,以研究跨系统发育和功能水平的变异,并评估趋同程度。
我们发现系统发育历史对颅骨和下颌骨形状有强烈影响,体型和基质仅起到次要作用。在形态空间中存在从较不专门化到更专门化的穴居者的明显梯度,沙中的穴居者与其他基质中的穴居者明显不同。我们还为四个物种创建了一个解剖学图谱,每个元素都单独描述。每个骨骼的形状和特征的相对比例都有一些变化,颅骨屋顶骨骼、筛骨、犁骨和腭骨变化最大。我们展示了与穴居性相关的特征的广泛趋同如何成为不同解剖变化的结果。
我们的研究使用几何形态测量学和比较解剖学来研究高度专业化和高要求的生活方式如何改变颅骨形态。尽管在形状和定性特征上有广泛的趋同,但系统发育历史起到了很大的作用,这种趋同很大程度上是由不同的解剖变化产生的,这意味着不同的发育途径或谱系特异性限制。即使在一个单一的科内,对专门化生态的适应也不会遵循单一的确定性路径。