Department of Medicinal Chemistry & Molecular Pharmacology, Purdue University, West Lafayette, Indiana, USA.
Department of Medicinal Chemistry & Molecular Pharmacology, Purdue University, West Lafayette, Indiana, USA; Department of Chemistry and Biochemistry, University of Notre Dame, Notre Dame, Indiana, USA.
J Biol Chem. 2021 Jan-Jun;296:100796. doi: 10.1016/j.jbc.2021.100796. Epub 2021 May 18.
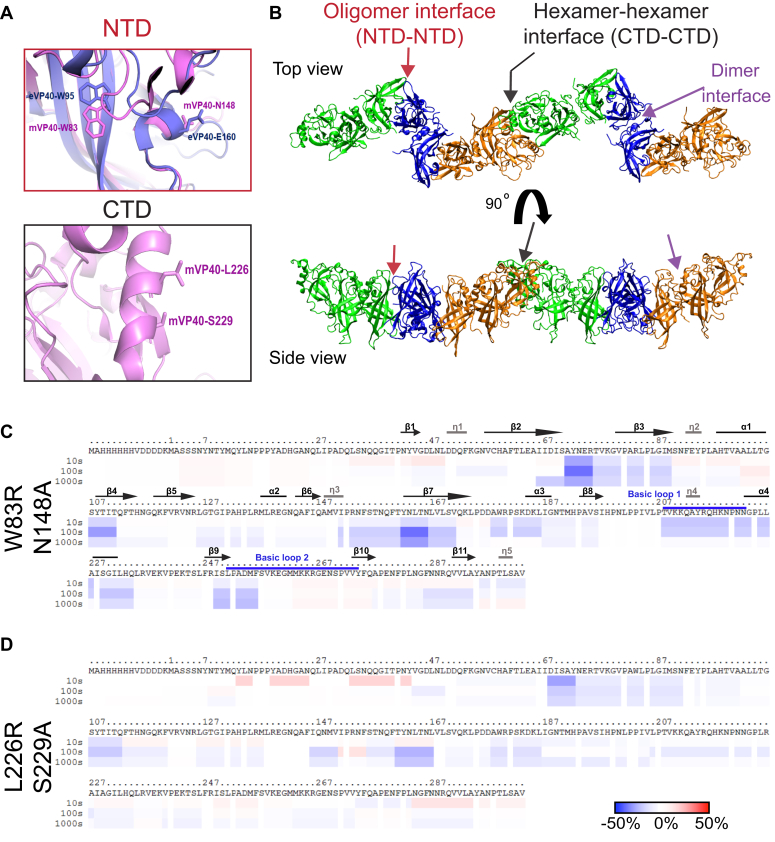
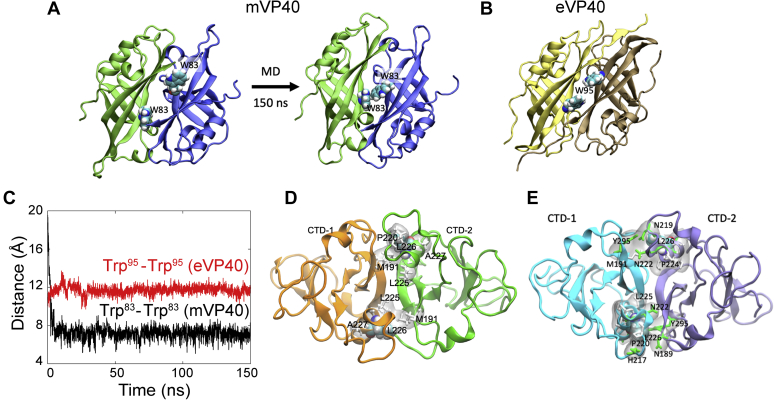
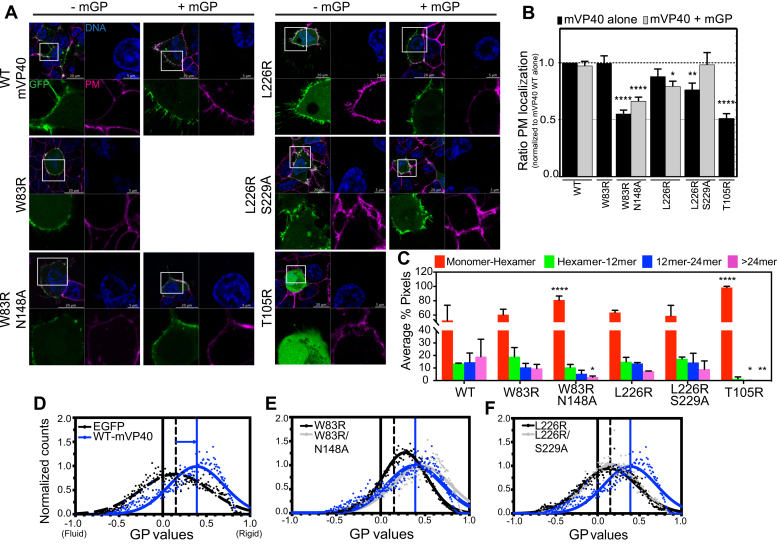
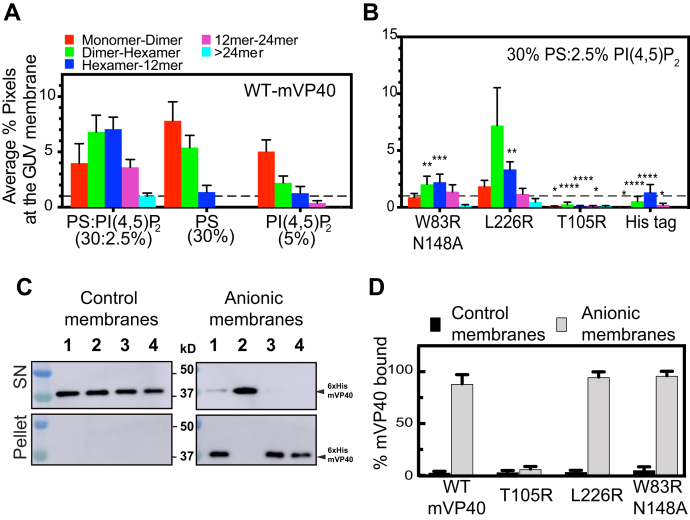
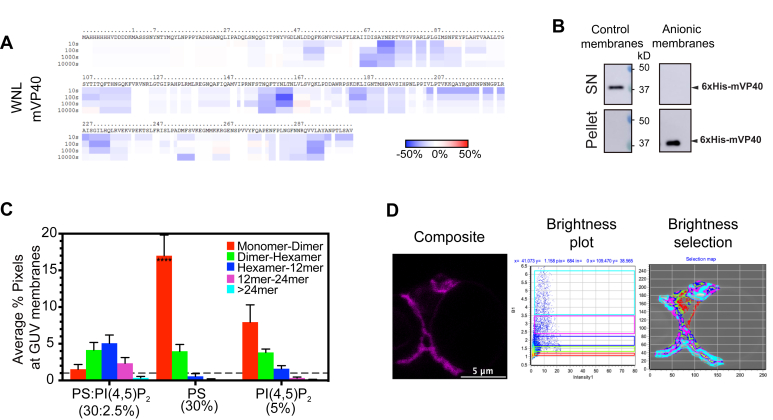
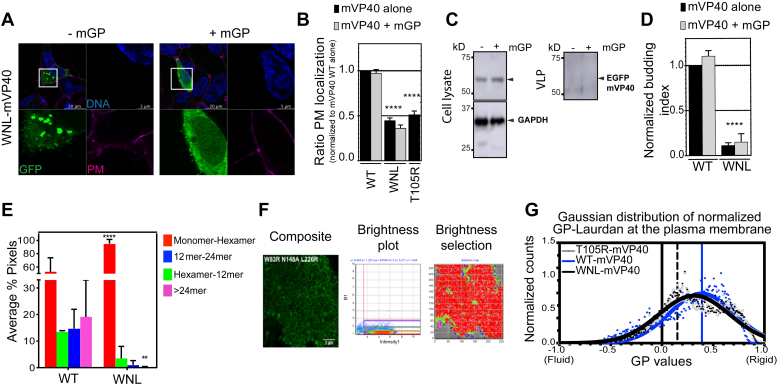
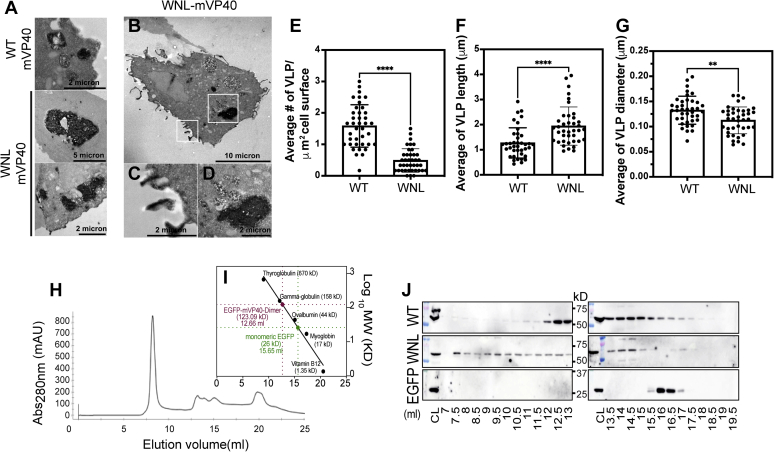
Marburg virus (MARV) is a lipid-enveloped virus harboring a negative-sense RNA genome, which has caused sporadic outbreaks of viral hemorrhagic fever in sub-Saharan Africa. MARV assembles and buds from the host cell plasma membrane where MARV matrix protein (mVP40) dimers associate with anionic lipids at the plasma membrane inner leaflet and undergo a dynamic and extensive self-oligomerization into the structural matrix layer. The MARV matrix layer confers the virion filamentous shape and stability but how host lipids modulate mVP40 oligomerization is mostly unknown. Using in vitro and cellular techniques, we present a mVP40 assembly model highlighting two distinct oligomerization interfaces: the (N-terminal domain [NTD] and C-terminal domain [CTD]) in mVP40. Cellular studies of NTD and CTD oligomerization interface mutants demonstrate the importance of each interface in matrix assembly. The assembly steps include protein trafficking to the plasma membrane, homo-multimerization that induced protein enrichment, plasma membrane fluidity changes, and elongations at the plasma membrane. An ascorbate peroxidase derivative (APEX)-transmission electron microscopy method was employed to closely assess the ultrastructural localization and formation of viral particles for wildtype mVP40 and NTD and CTD oligomerization interface mutants. Taken together, these studies present a mechanistic model of mVP40 oligomerization and assembly at the plasma membrane during virion assembly that requires interactions with phosphatidylserine for NTD-NTD interactions and phosphatidylinositol-4,5-bisphosphate for proper CTD-CTD interactions. These findings have broader implications in understanding budding of lipid-enveloped viruses from the host cell plasma membrane and potential strategies to target protein-protein or lipid-protein interactions to inhibit virus budding.
马尔堡病毒(MARV)是一种带有负义 RNA 基因组的脂包膜病毒,已在撒哈拉以南非洲地区引起散发性病毒性出血热暴发。MARV 在宿主细胞膜的质膜中组装和出芽,其中 MARV 基质蛋白(mVP40)二聚体与质膜内层的阴离子脂质结合,并在质膜中动态且广泛地自我寡聚化为结构基质层。MARV 基质层赋予病毒丝状形状和稳定性,但宿主脂质如何调节 mVP40 寡聚化在很大程度上尚不清楚。我们使用体外和细胞技术提出了一个 mVP40 组装模型,突出了两个不同的寡聚化界面:mVP40 的(N 端结构域 [NTD] 和 C 端结构域 [CTD])。对 NTD 和 CTD 寡聚化界面突变体的细胞研究表明了每个界面在基质组装中的重要性。组装步骤包括蛋白质向质膜的转运、诱导蛋白富集的同源多聚化、质膜流动性的变化以及在质膜上的伸长。我们采用抗坏血酸过氧化物酶衍生物(APEX)-透射电子显微镜方法密切评估野生型 mVP40 和 NTD 和 CTD 寡聚化界面突变体的病毒颗粒的超微结构定位和形成。总之,这些研究提出了一个在病毒组装过程中 mVP40 在质膜上寡聚化和组装的机制模型,该模型需要与质膜内层的磷脂酰丝氨酸相互作用以进行 NTD-NTD 相互作用,以及与磷脂酰肌醇-4,5-二磷酸相互作用以进行正确的 CTD-CTD 相互作用。这些发现对于理解从宿主细胞膜出芽的脂质包膜病毒以及潜在的靶向蛋白-蛋白或脂质-蛋白相互作用以抑制病毒出芽的策略具有更广泛的意义。