Centers for Cardiovascular Research, Lewis Katz School of Medicine at Temple University, Philadelphia, PA, United States.
Metabolic Disease Research & Thrombosis Research, Department of Cardiovascular Sciences, Lewis Katz School of Medicine at Temple University, Philadelphia, PA, United States.
Front Immunol. 2021 May 18;12:678201. doi: 10.3389/fimmu.2021.678201. eCollection 2021.
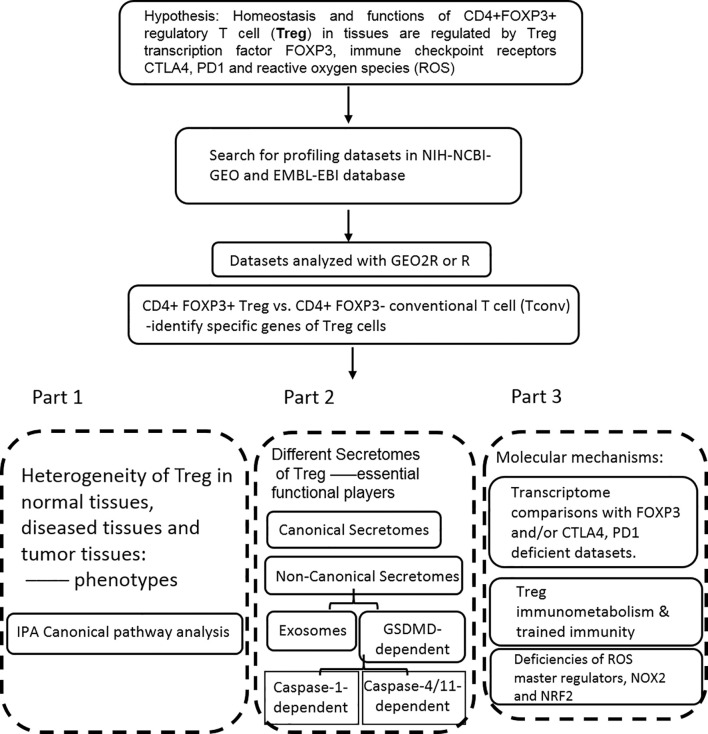
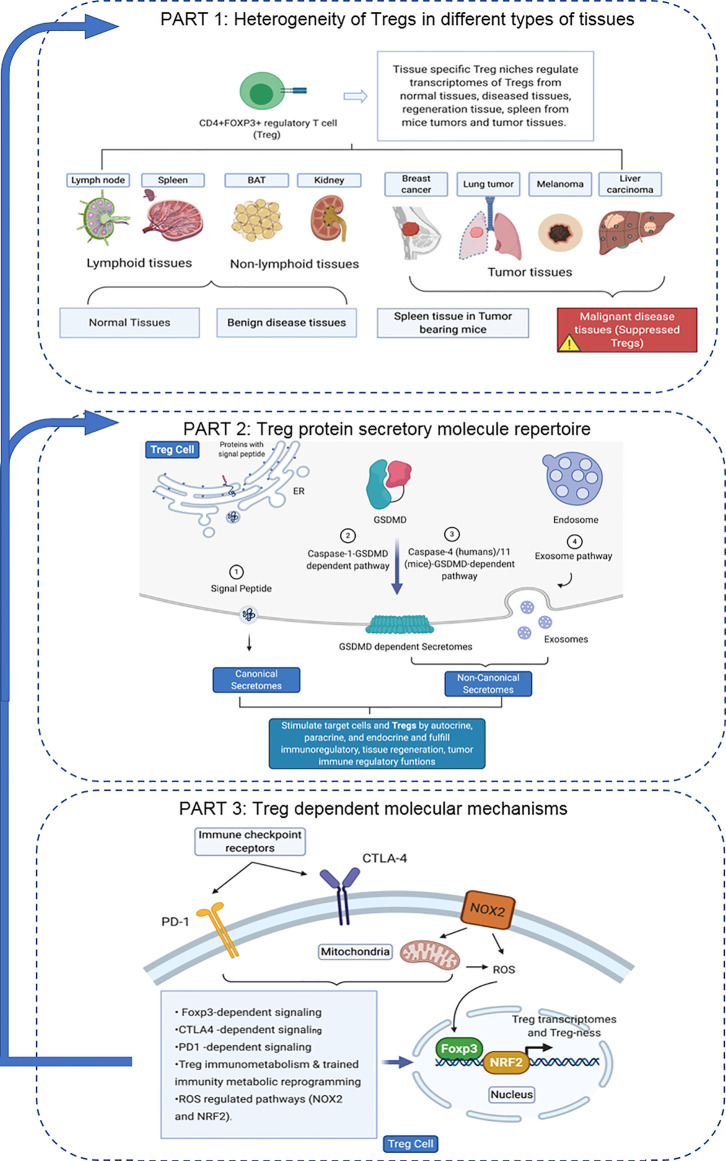
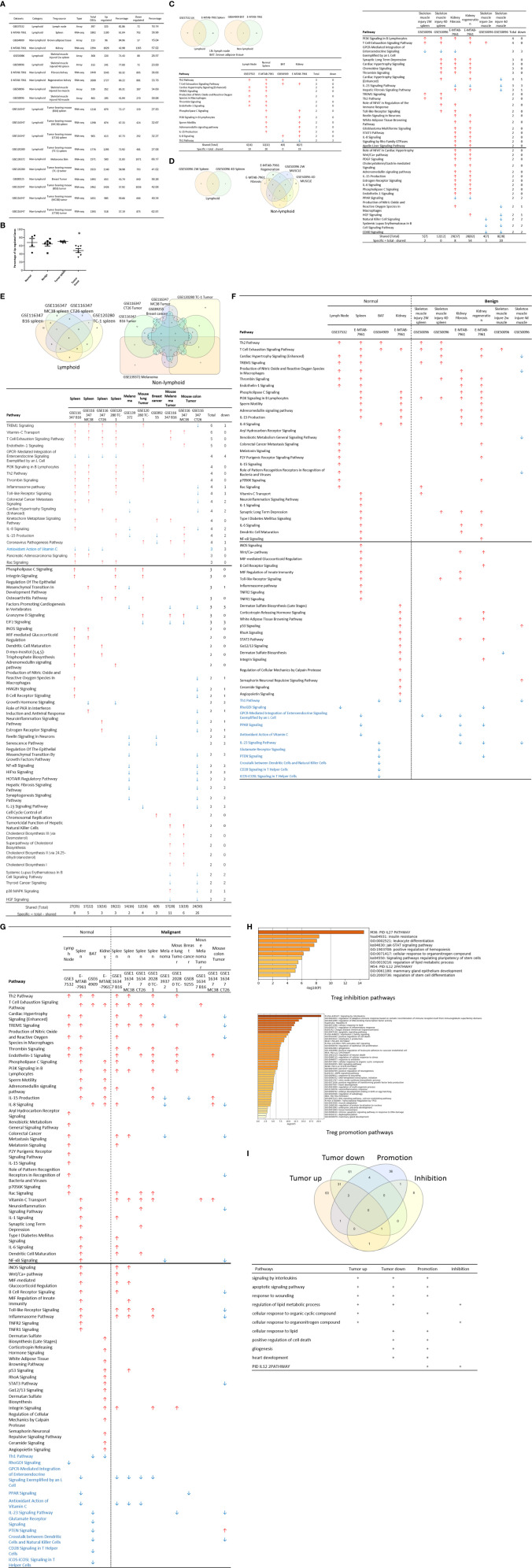
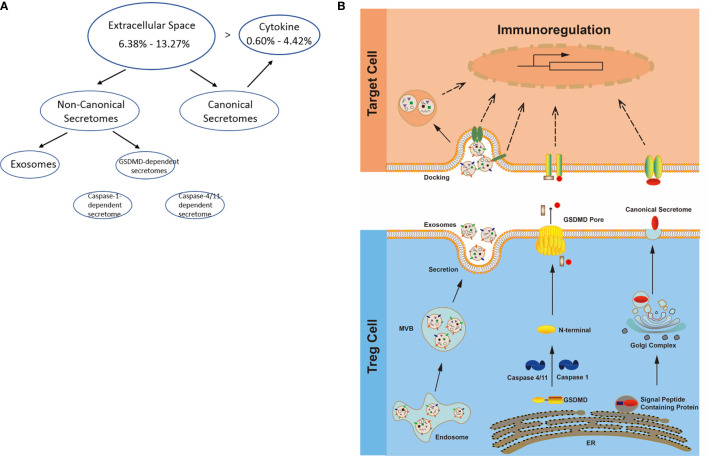
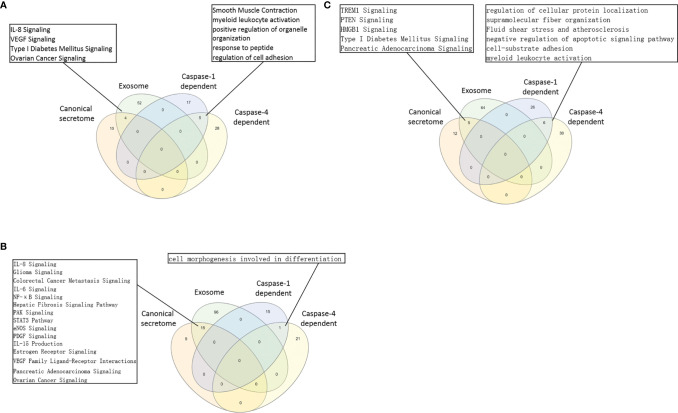
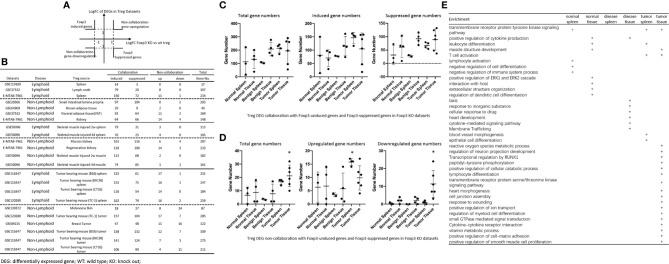
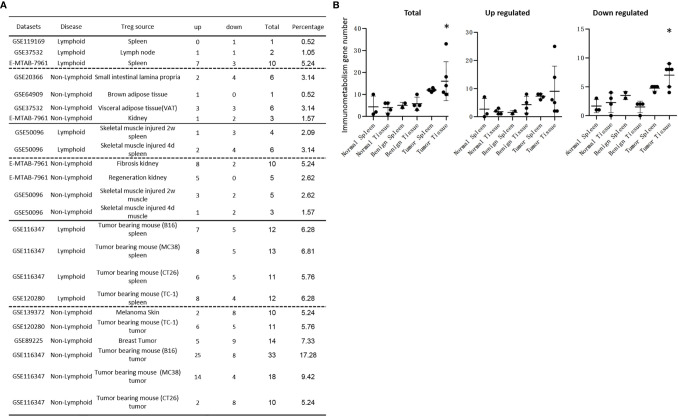
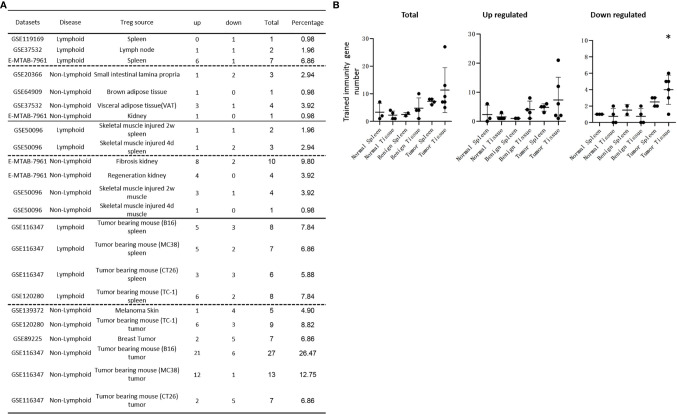
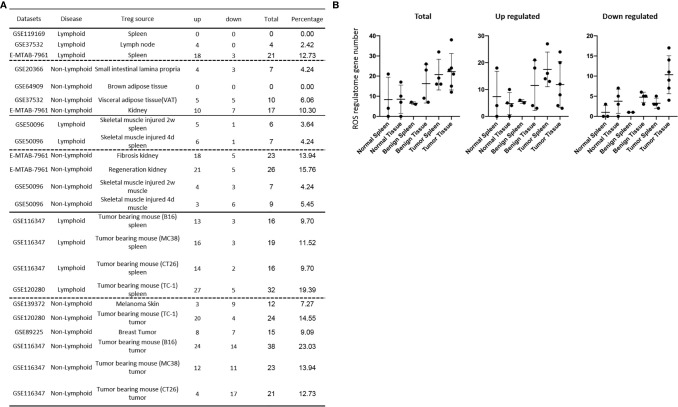
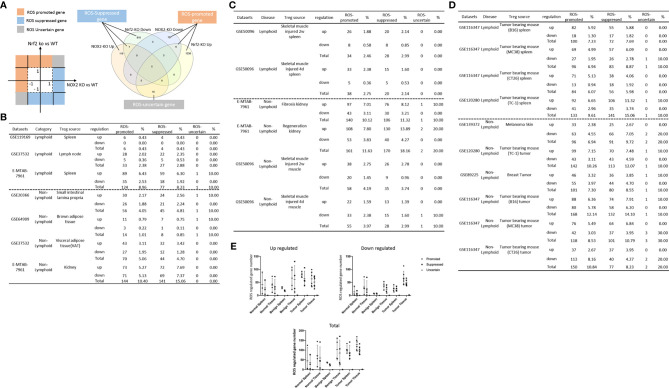
We performed a transcriptomic analyses using the strategies we pioneered and made the following findings: Normal lymphoid Tregs, diseased kidney Tregs, splenic Tregs from mice with injured muscle have 3, 17 and 3 specific (S-) pathways, respectively; Tumor splenic Tregs share 12 pathways with tumor Tregs; tumor splenic Tregs and tumor Tregs have 11 and 8 S-pathways, respectively; Normal and non-tumor disease Tregs upregulate some of novel 2641 canonical secretomic genes (SGs) with 24 pathways, and tumor Tregs upregulate canonical secretomes with 17 pathways; 4) Normal and non-tumor disease tissue Tregs upregulate some of novel 6560 exosome SGs with 56 exosome SG pathways (ESP), tumor Treg ESP are more focused than other Tregs; 5) Normal, non-tumor diseased Treg and tumor Tregs upregulate some of novel 961 innate immune caspase-1 SGs and 1223 innate immune caspase-4 SGs to fulfill their tissue/SG-specific and shared functions; Most tissue Treg transcriptomes are controlled by Foxp3; and Tumor Tregs had increased Foxp3 non-collaboration genes with ROS and 17 other pathways; Immune checkpoint receptor PD-1 does, but CTLA-4 does not, play significant roles in promoting Treg upregulated genes in normal and non-tumor disease tissue Tregs; and tumor splenic and tumor Tregs have certain CTLA-4-, and PD-1-, non-collaboration transcriptomic changes with innate immune dominant pathways; Tumor Tregs downregulate more immunometabolic and innate immune memory (trained immunity) genes than Tregs from other groups; and ROS significantly regulate Treg transcriptomes; and ROS-suppressed genes are downregulated more in tumor Tregs than Tregs from other groups. Our results have provided novel insights on the roles of Tregs in normal, injuries, regeneration, tumor conditions and some of canonical and innate immune non-canonical secretomes ROS-regulatory mechanisms and new therapeutic targets for immunosuppression, tissue repair, cardiovascular diseases, chronic kidney disease, autoimmune diseases, transplantation, and cancers.
我们使用我们首创的转录组分析策略进行了研究,得出以下发现:正常淋巴样 Tregs、患病肾脏 Tregs、受损肌肉小鼠脾脏 Tregs 分别具有 3、17 和 3 个特异性(S-)途径;肿瘤脾脏 Tregs 与肿瘤 Tregs 有 12 个途径共享;肿瘤脾脏 Tregs 和肿瘤 Tregs 分别有 11 和 8 个 S-途径;正常和非肿瘤疾病 Tregs 上调了一些具有 24 条途径的新型 2641 个经典分泌组基因(SGs),肿瘤 Tregs 上调了具有 17 条途径的经典分泌组;正常和非肿瘤疾病组织 Tregs 上调了一些具有 56 个外泌体 SG 途径(ESP)的新型 6560 个外泌体 SGs,肿瘤 Treg ESP 比其他 Tregs 更集中;正常、非肿瘤疾病 Treg 和肿瘤 Tregs 上调了一些新型的先天免疫 Caspase-1 和 Caspase-4 SGs,以发挥其组织/ SG 特异性和共享功能;大多数组织 Treg 转录组受 Foxp3 控制;肿瘤 Tregs 增加了与 ROS 和 17 条其他途径非协作的基因;免疫检查点受体 PD-1 确实但 CTLA-4 没有在促进正常和非肿瘤疾病组织 Tregs 上调基因中发挥重要作用;肿瘤脾脏和肿瘤 Tregs 具有某些 CTLA-4-和 PD-1-非协作转录组变化与先天免疫主导途径;肿瘤 Tregs 下调的免疫代谢和先天免疫记忆(训练免疫)基因多于其他组的 Tregs;ROS 显著调节 Treg 转录组;ROS 抑制基因在肿瘤 Tregs 中的下调程度高于其他组。我们的研究结果为 Tregs 在正常、损伤、再生、肿瘤状态以及一些经典和先天免疫非经典分泌组中的作用提供了新的见解 ROS 调节机制和新的免疫抑制、组织修复、心血管疾病、慢性肾脏病、自身免疫性疾病、移植和癌症的治疗靶点。