Hunter James Kelly Research Institute, University at Buffalo, Buffalo, United States.
Department of Biochemistry, University at Buffalo, Buffalo, United States.
Elife. 2021 Sep 14;10:e66278. doi: 10.7554/eLife.66278.
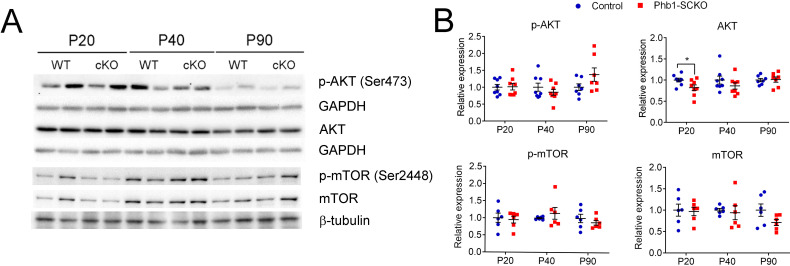
Schwann cell (SC) mitochondria are quickly emerging as an important regulator of myelin maintenance in the peripheral nervous system (PNS). However, the mechanisms underlying demyelination in the context of mitochondrial dysfunction in the PNS are incompletely understood. We recently showed that conditional ablation of the mitochondrial protein Prohibitin 1 (PHB1) in SCs causes a severe and fast progressing demyelinating peripheral neuropathy in mice, but the mechanism that causes failure of myelin maintenance remained unknown. Here, we report that mTORC1 and c-Jun are continuously activated in the absence of , likely as part of the SC response to mitochondrial damage. Moreover, we demonstrate that these pathways are involved in the demyelination process, and that inhibition of mTORC1 using rapamycin partially rescues the demyelinating pathology. Therefore, we propose that mTORC1 and c-Jun may play a critical role as executioners of demyelination in the context of perturbations to SC mitochondria.
施旺细胞(SC)的线粒体正在迅速成为外周神经系统(PNS)中髓鞘维持的一个重要调节因子。然而,在 PNS 中线粒体功能障碍的背景下脱髓鞘的机制还不完全清楚。我们最近表明,条件性敲除施旺细胞中线粒体蛋白 Prohibitin 1(PHB1)会导致小鼠严重且快速进展的脱髓鞘周围神经病,但导致髓鞘维持失败的机制仍不清楚。在这里,我们报告说,mTORC1 和 c-Jun 在 PHB1 缺失的情况下持续激活,这可能是 SC 对线粒体损伤反应的一部分。此外,我们证明这些途径参与了脱髓鞘过程,并且使用雷帕霉素抑制 mTORC1 可部分挽救脱髓鞘病理。因此,我们提出 mTORC1 和 c-Jun 可能作为 SC 线粒体扰动时脱髓鞘的执行者发挥关键作用。