Department of Anesthesiology and Critical Care Medicine, Johns Hopkins University, Baltimore, Maryland, USA.
Compr Physiol. 2021 Sep 23;11(4):2371-2432. doi: 10.1002/cphy.c200028.
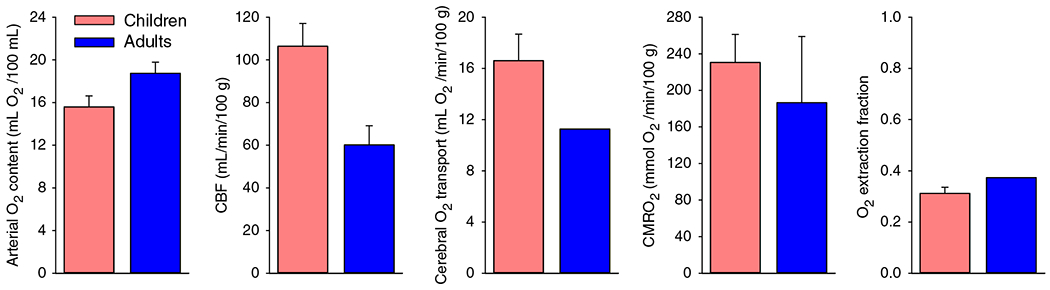
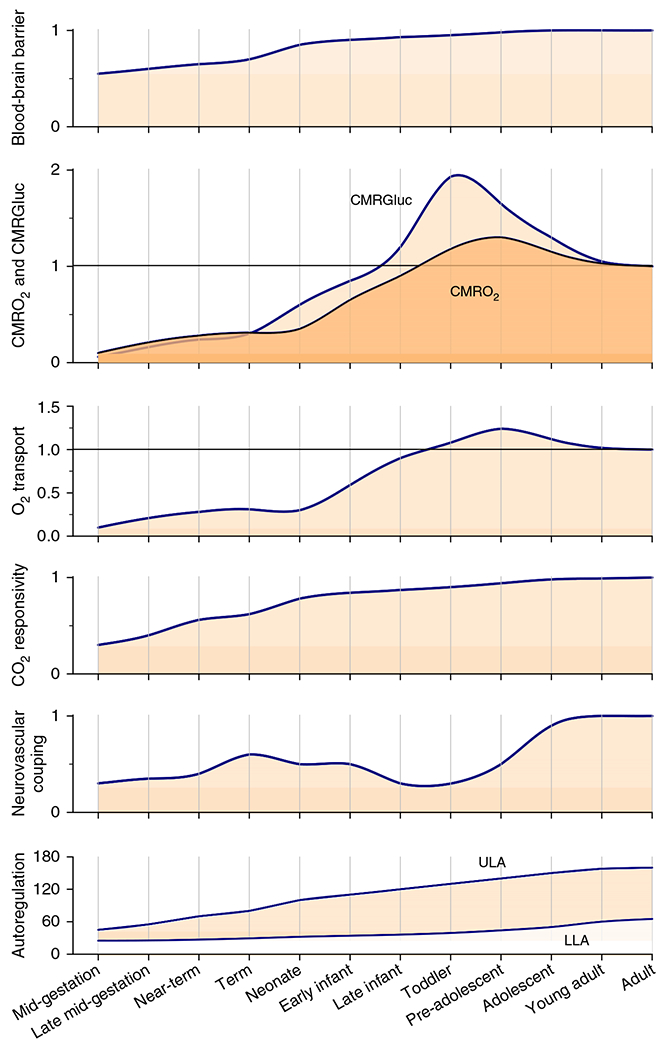
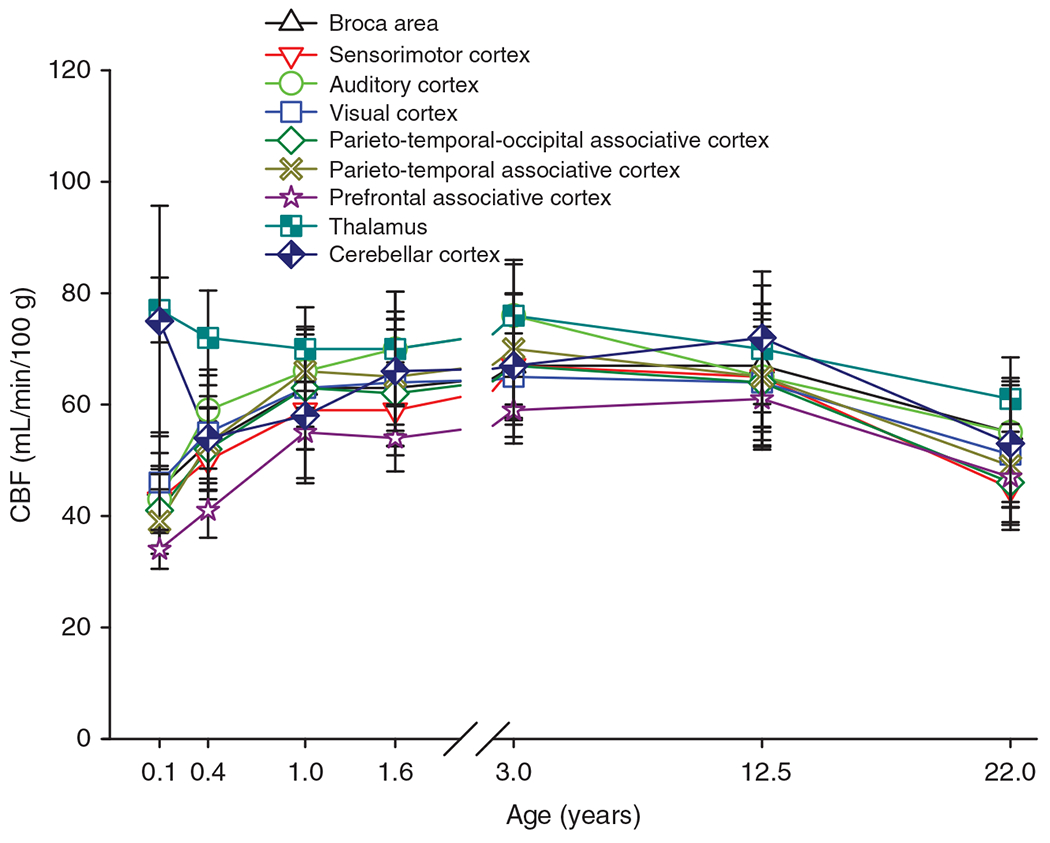
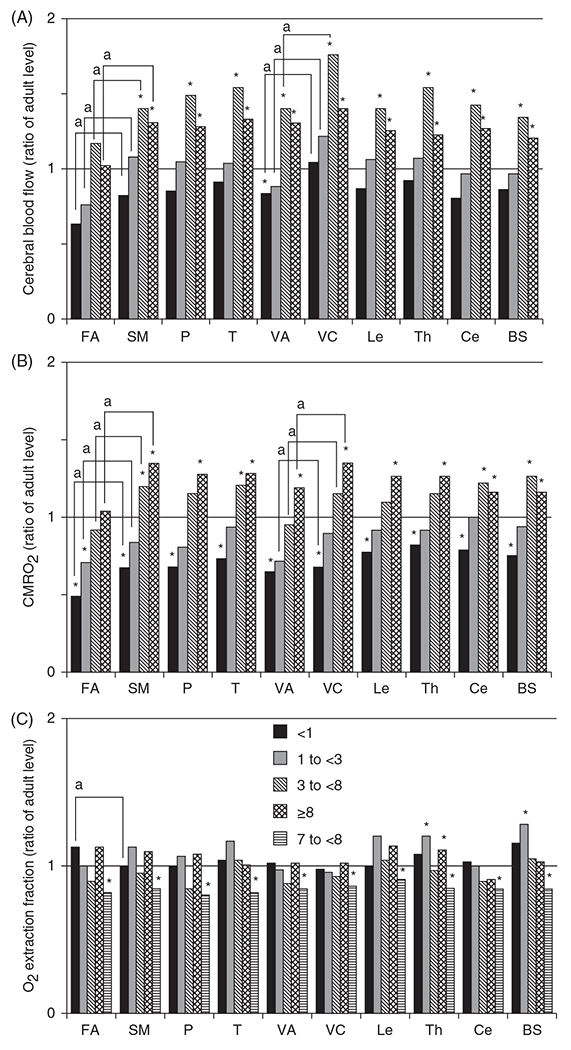
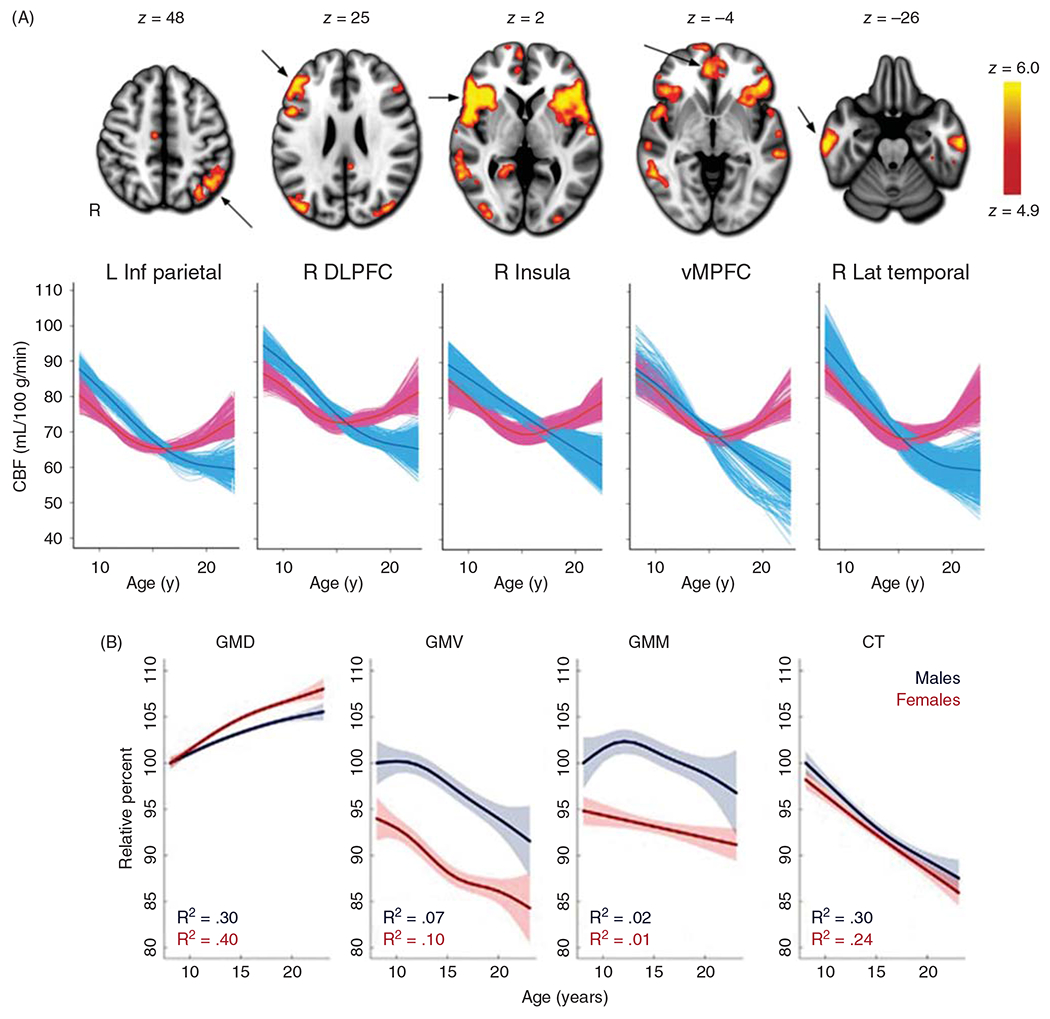
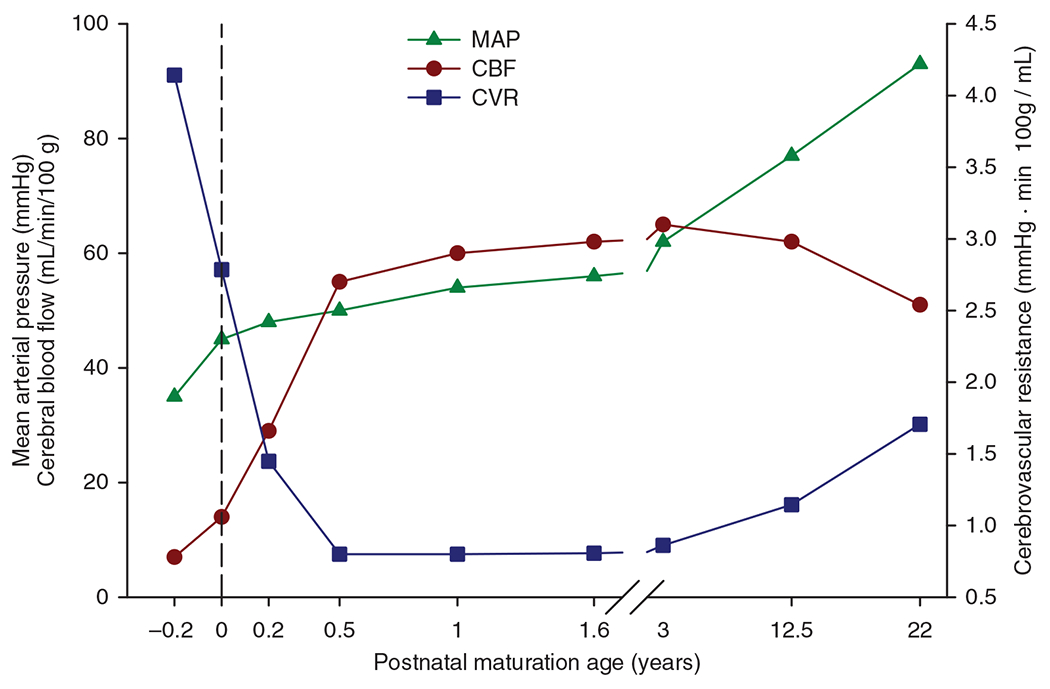
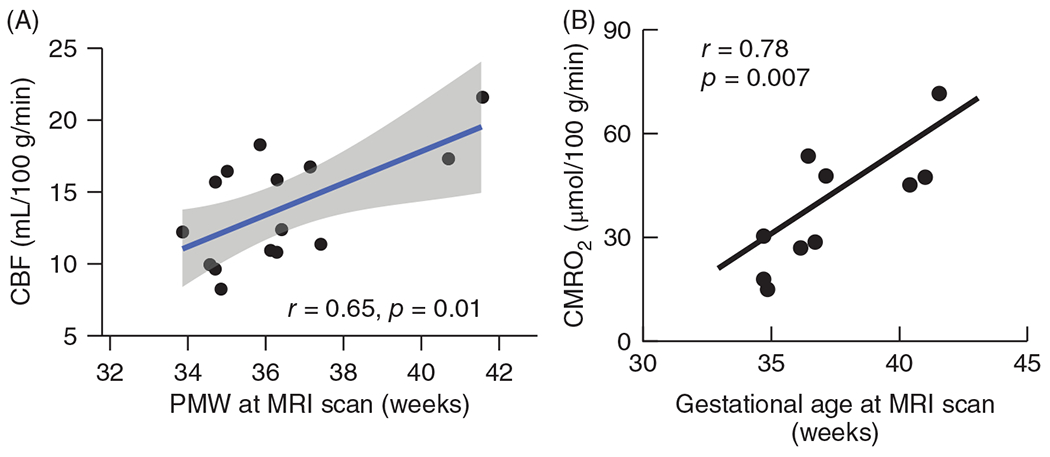
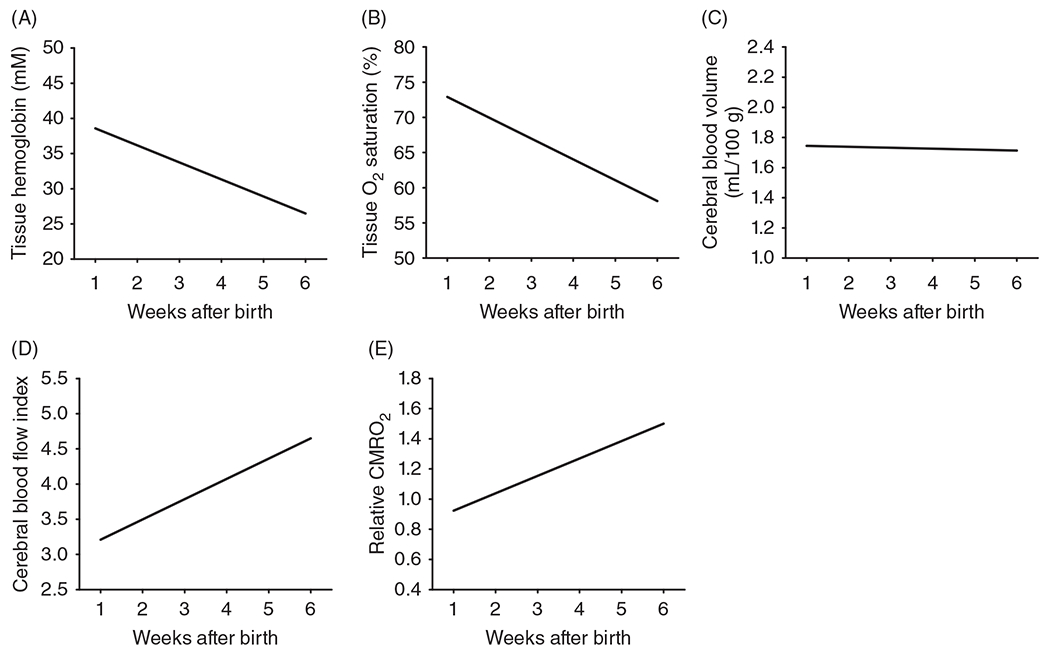
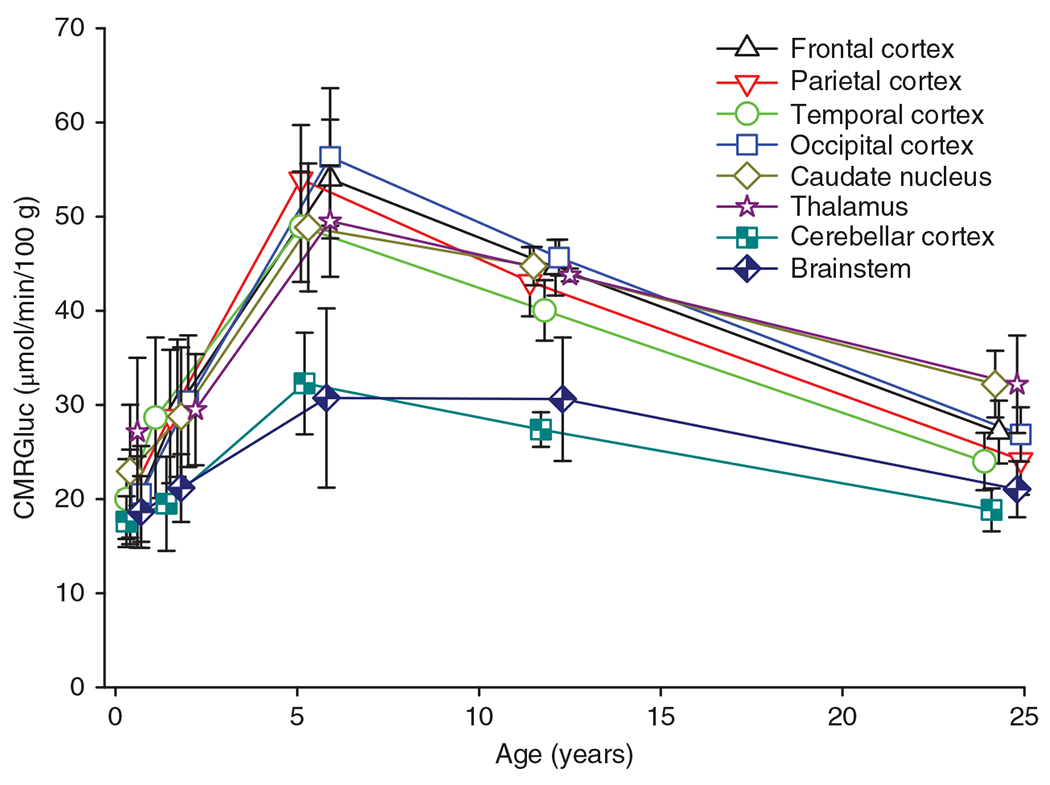
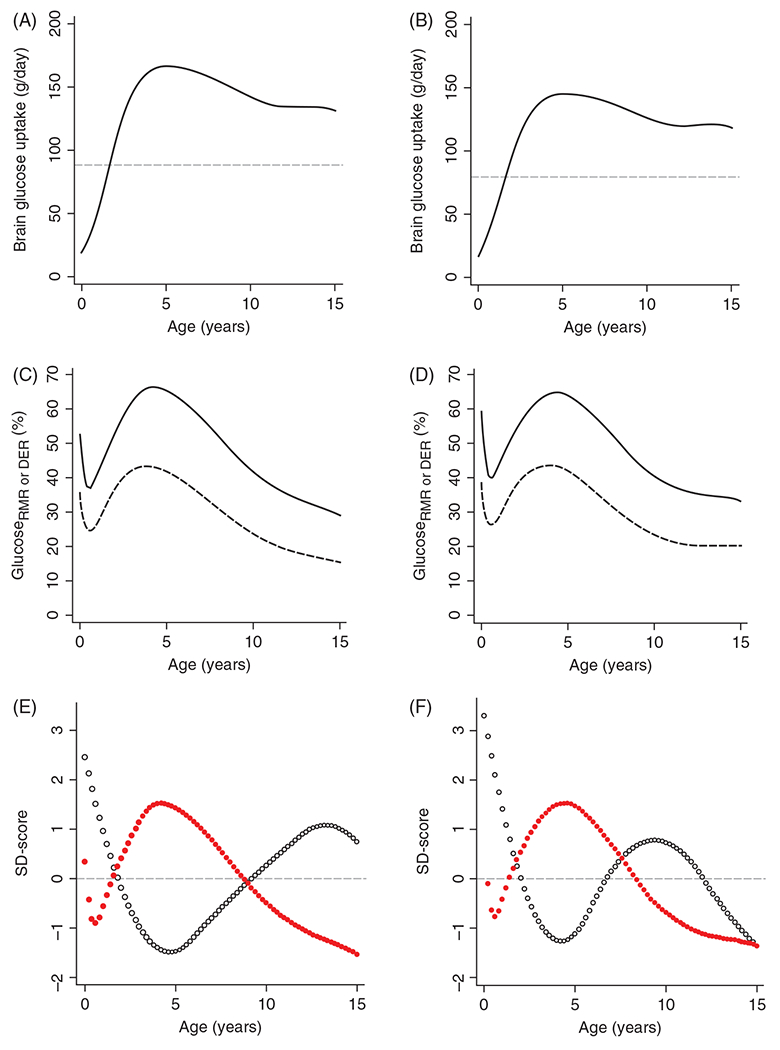
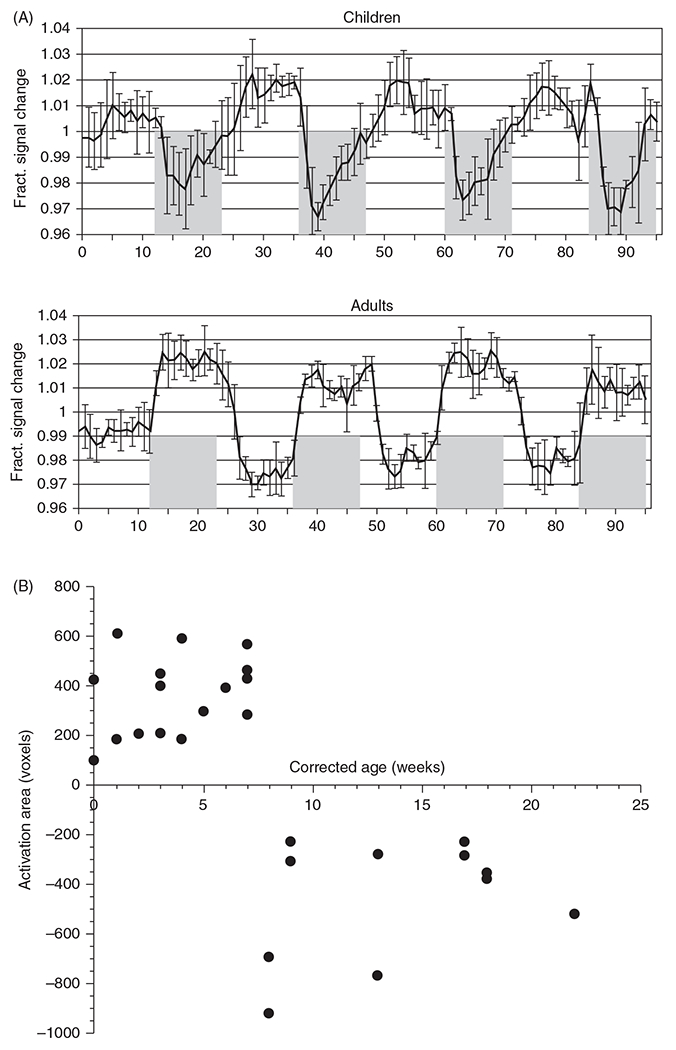
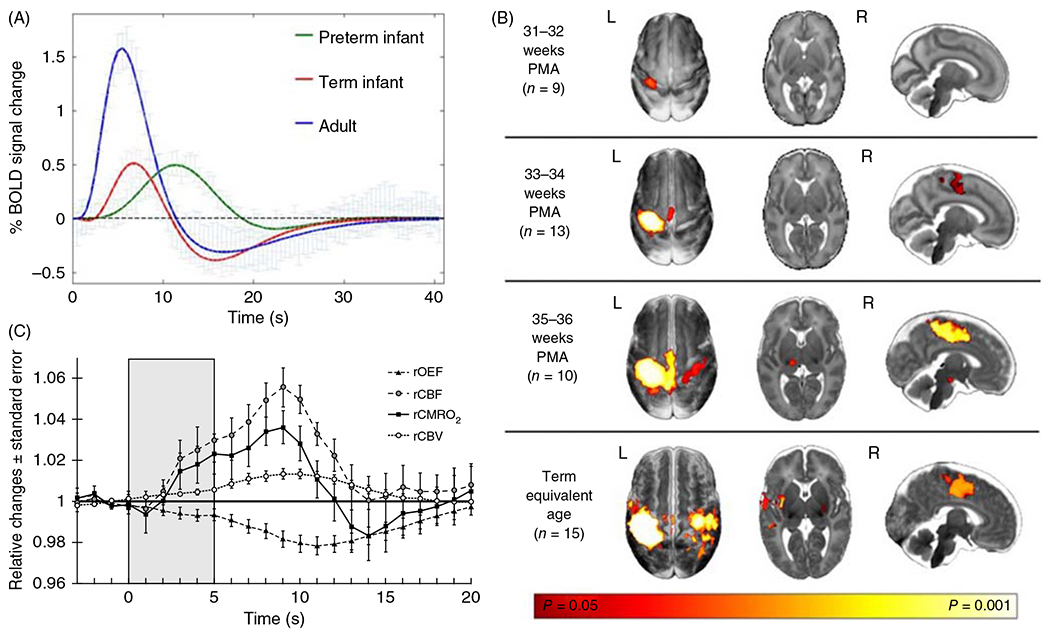
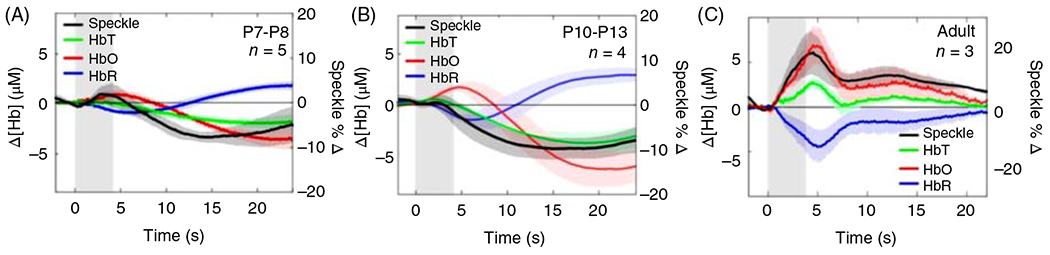
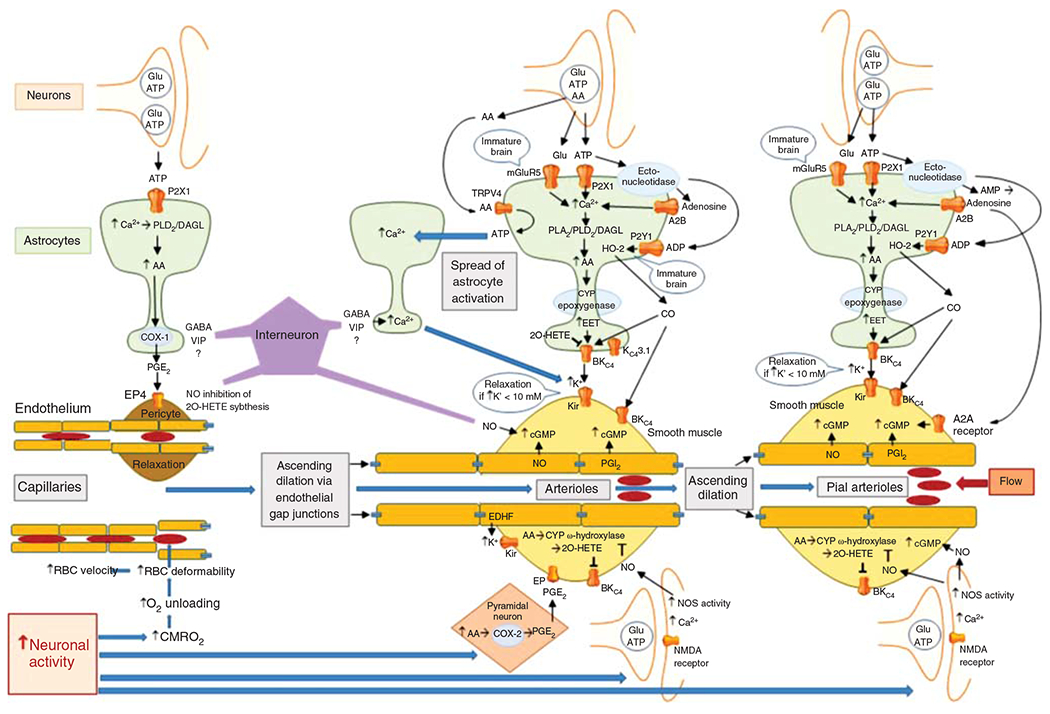
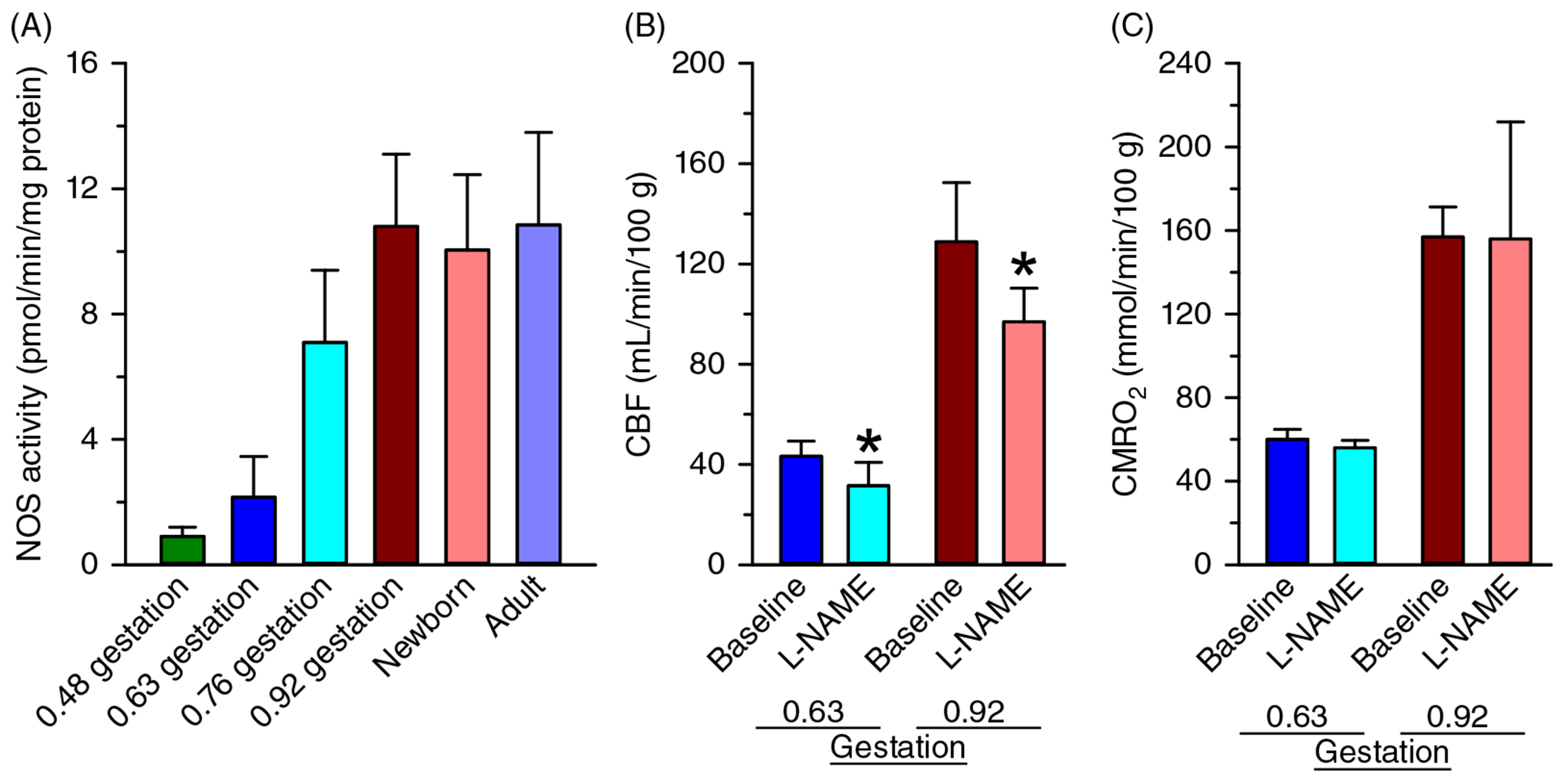
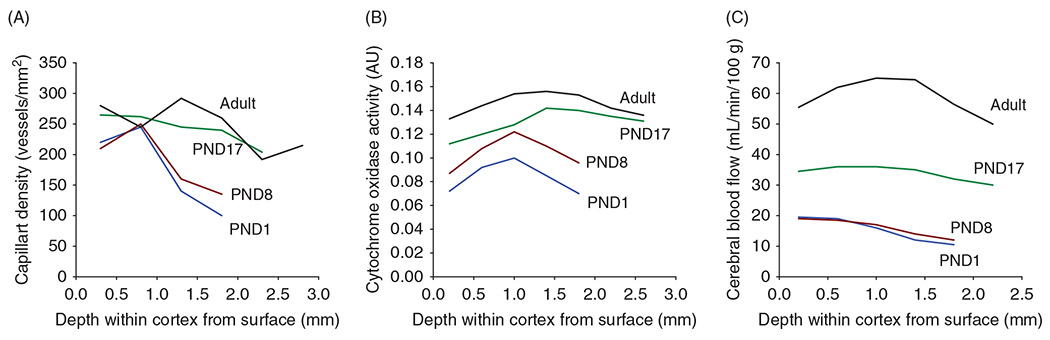
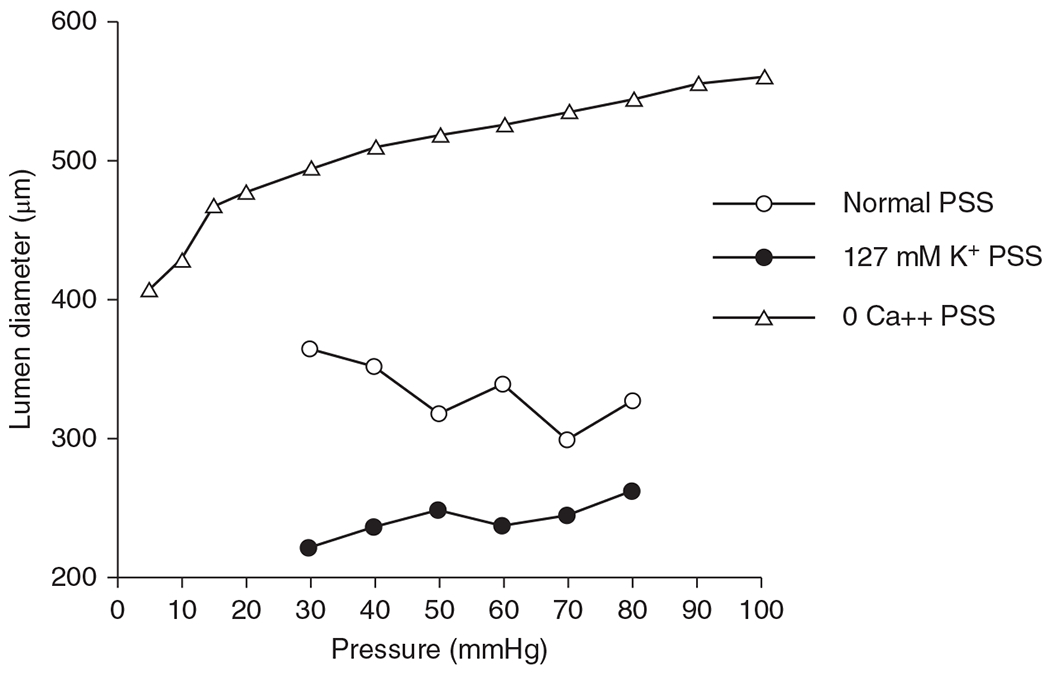
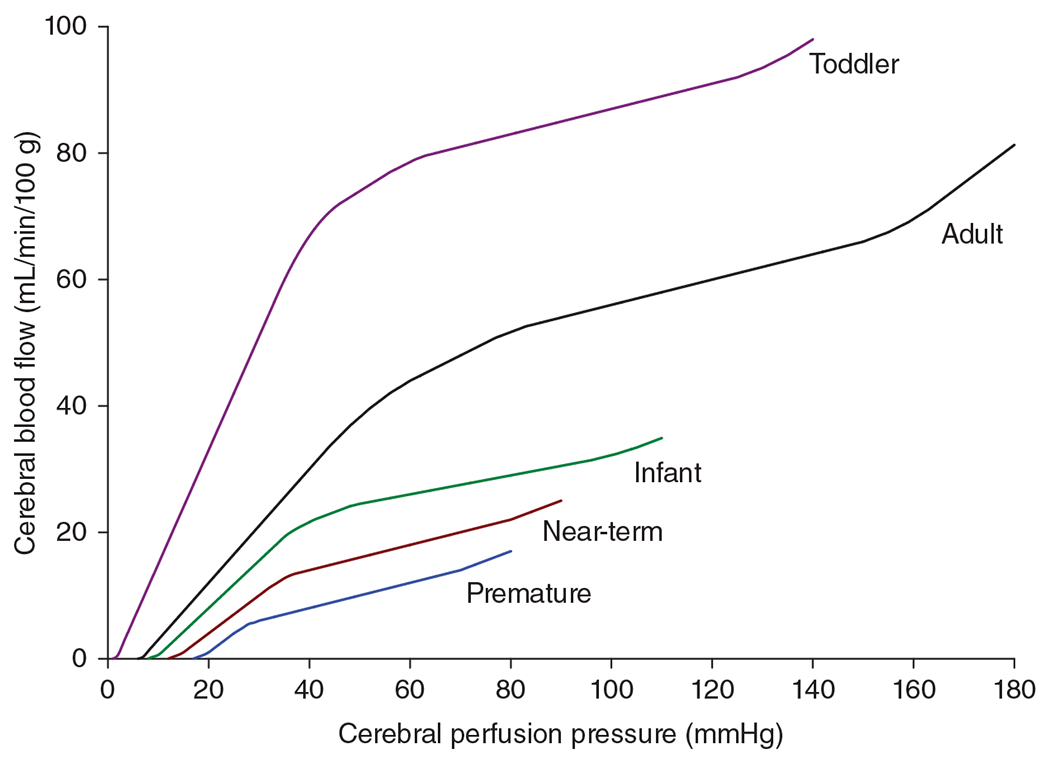
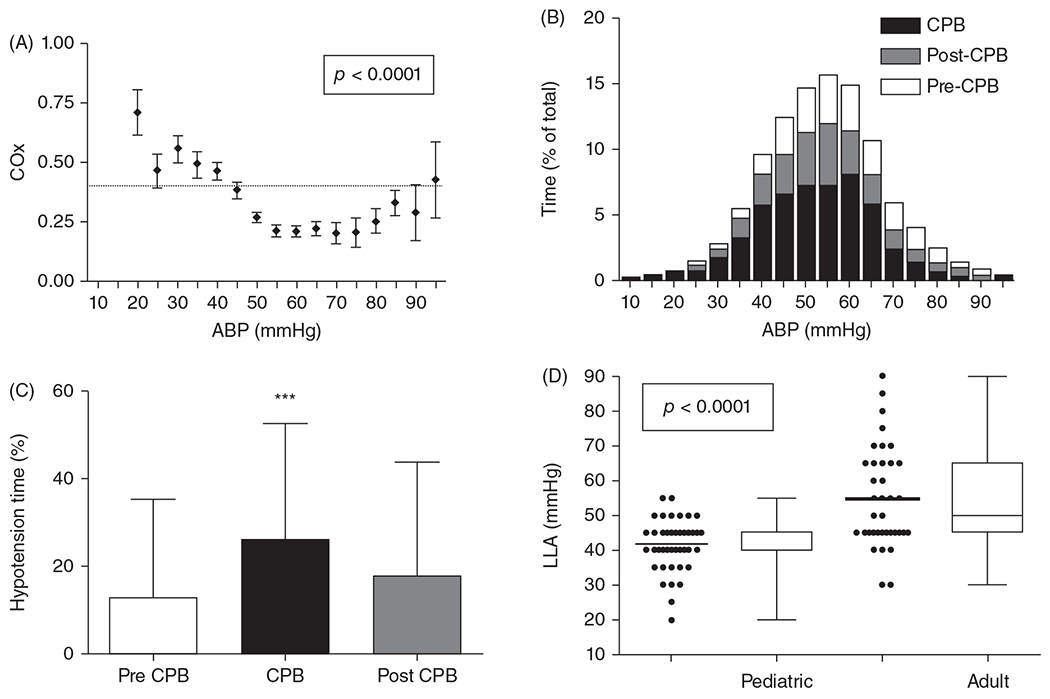
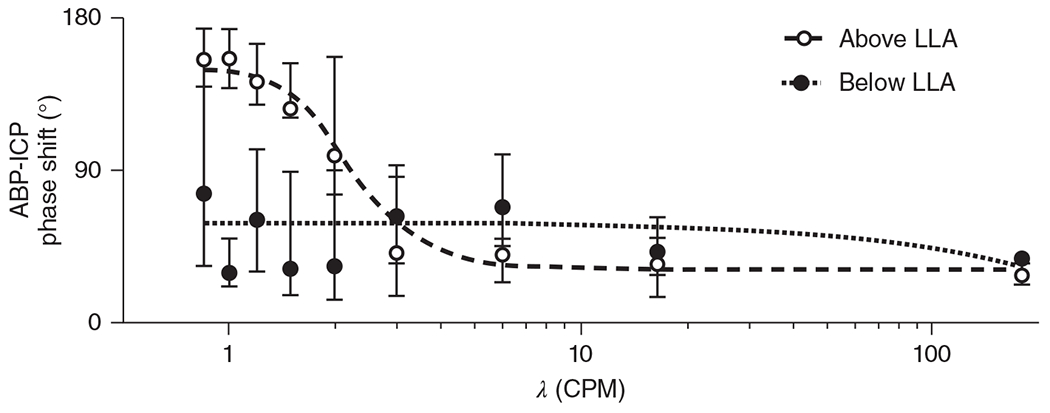
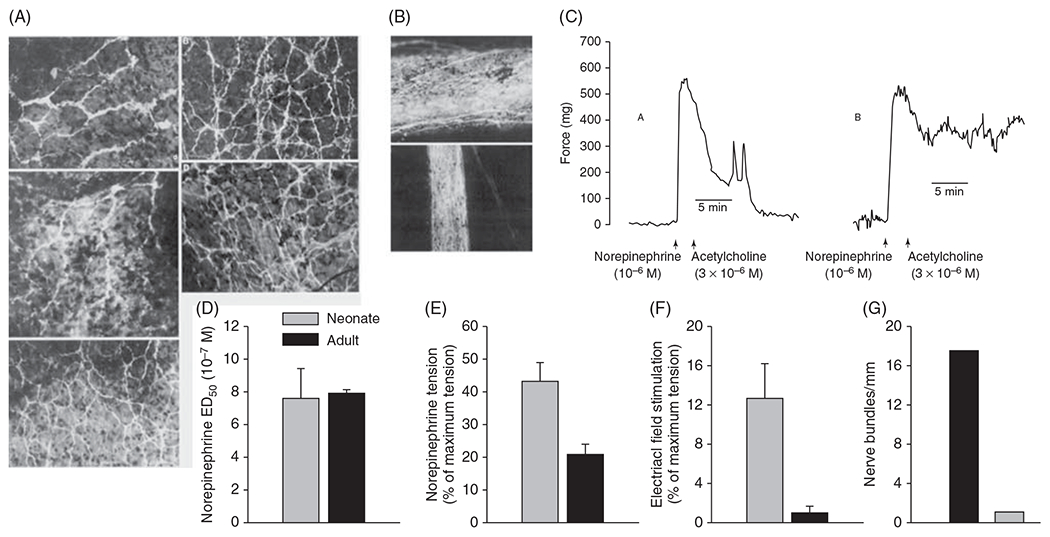
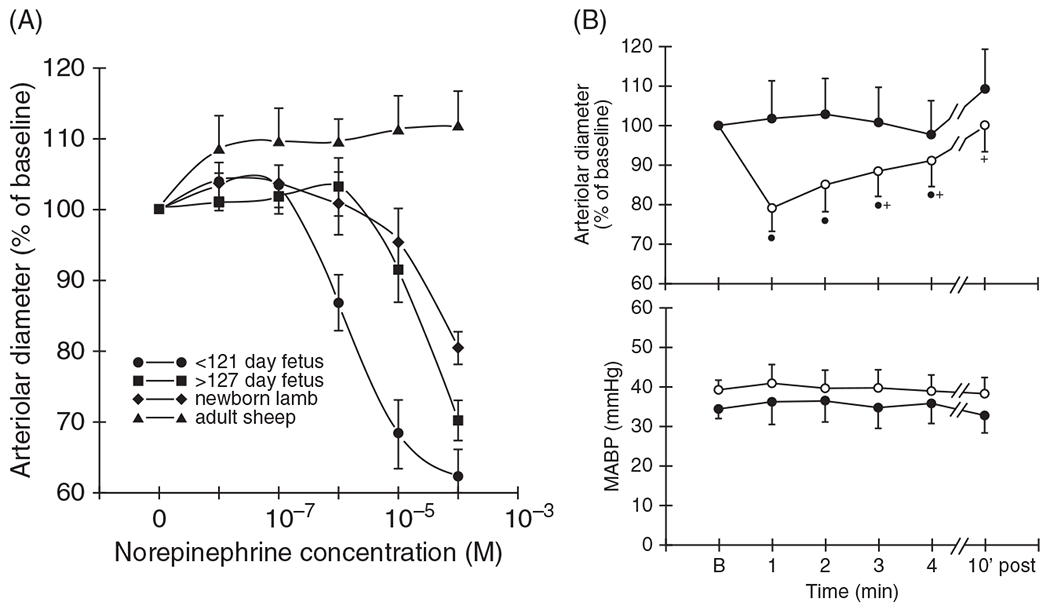
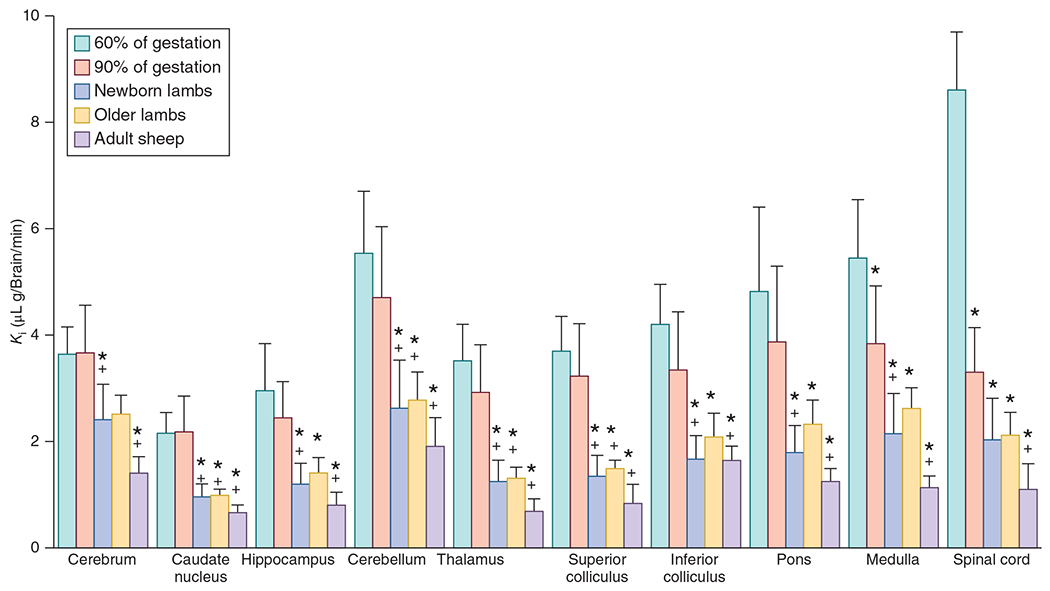
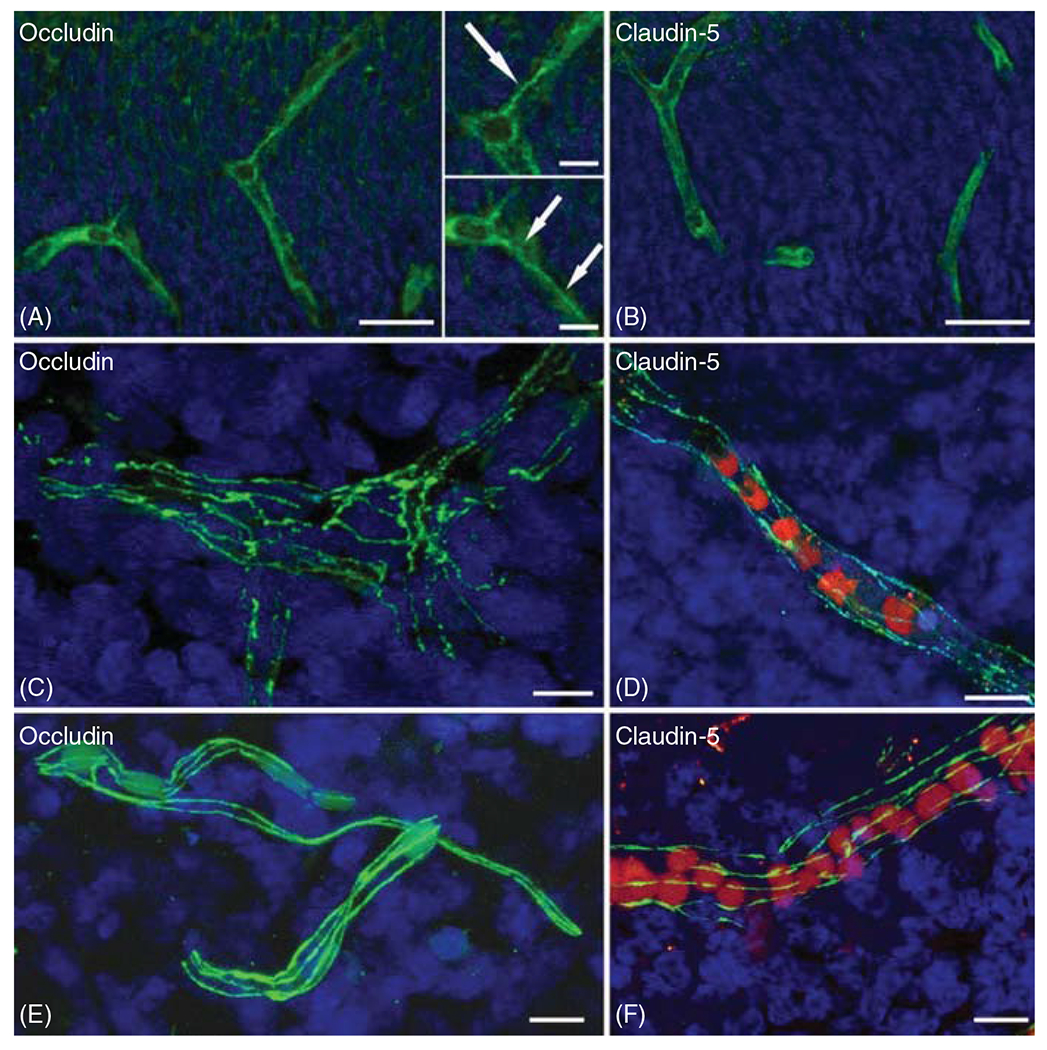
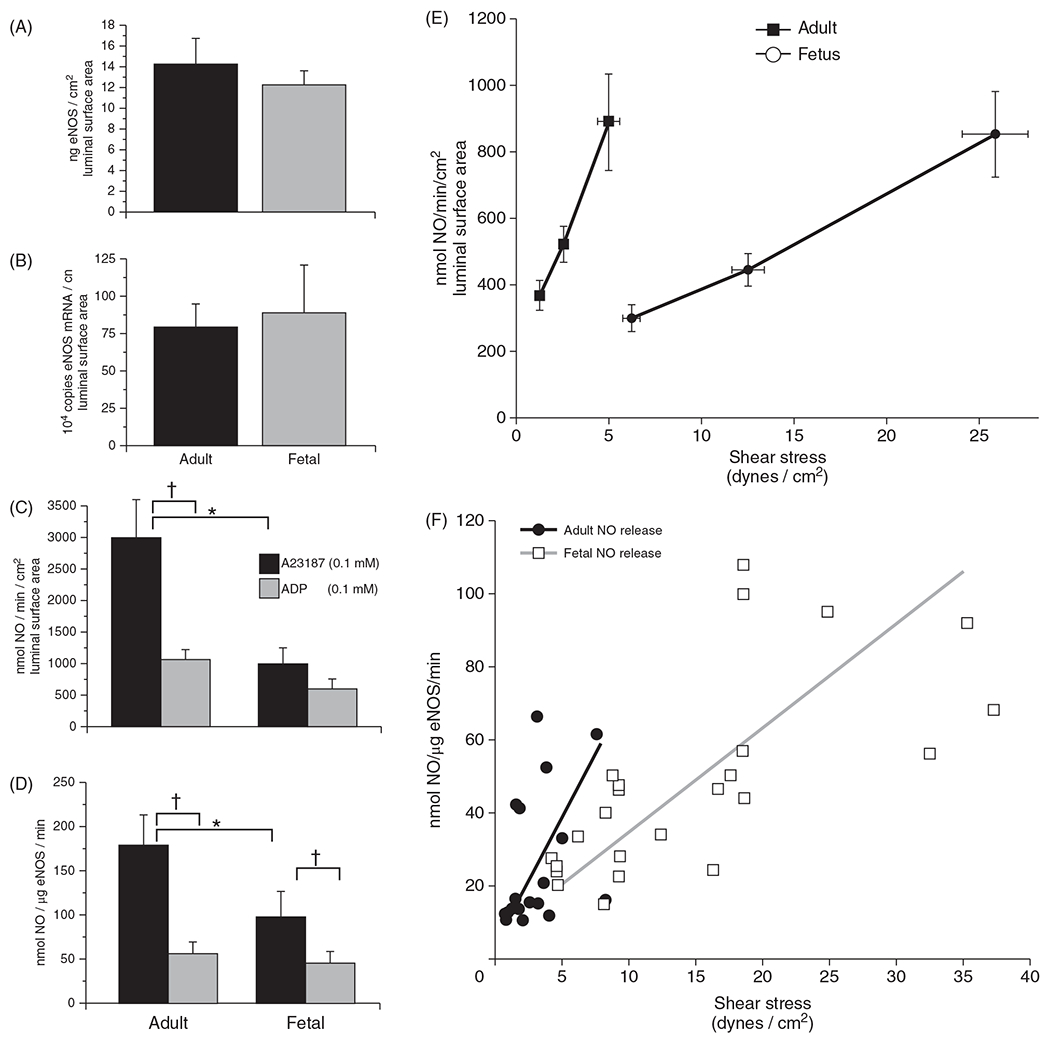
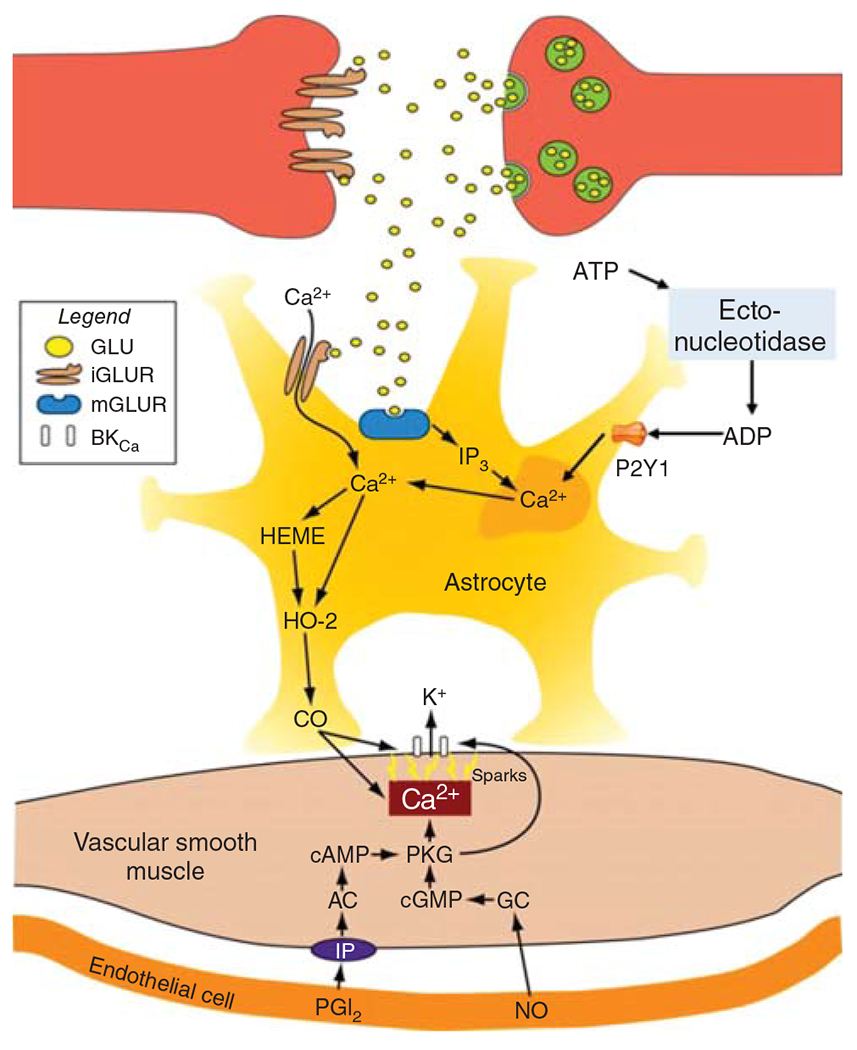
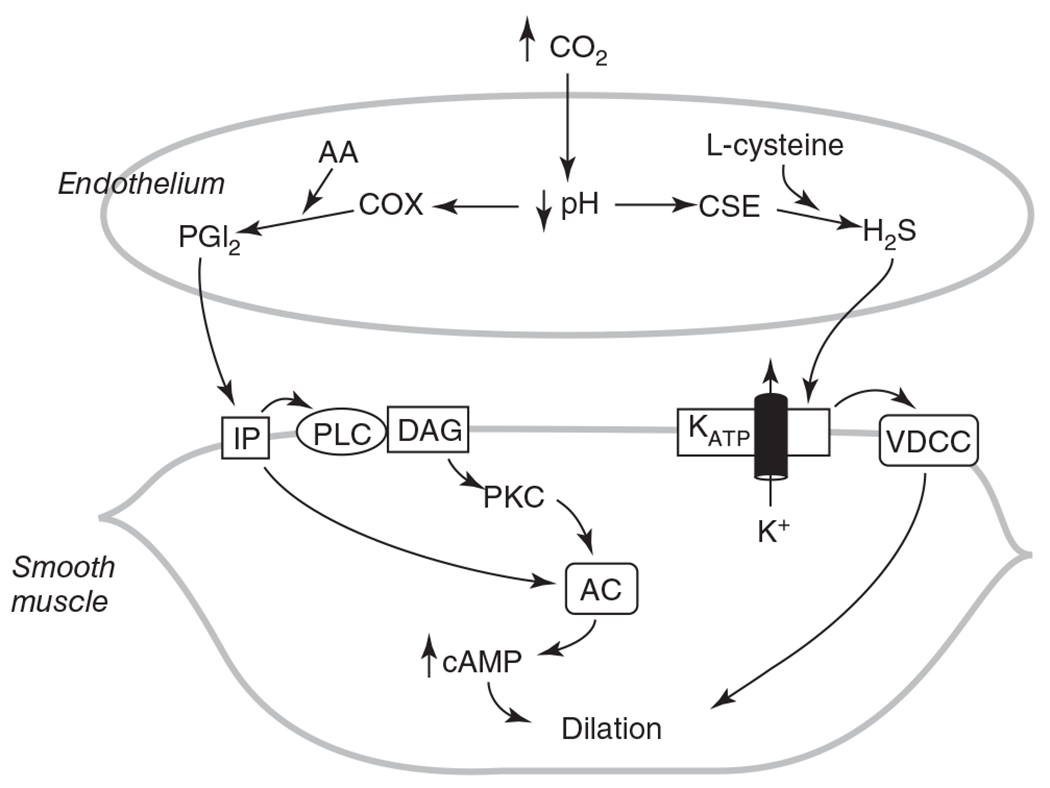
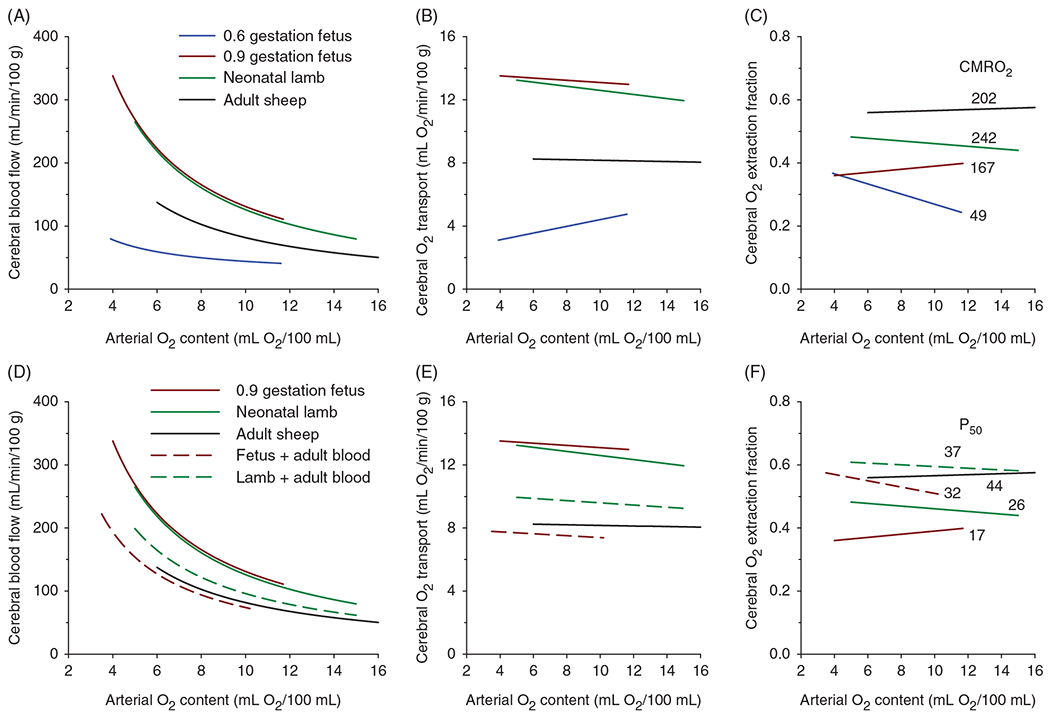
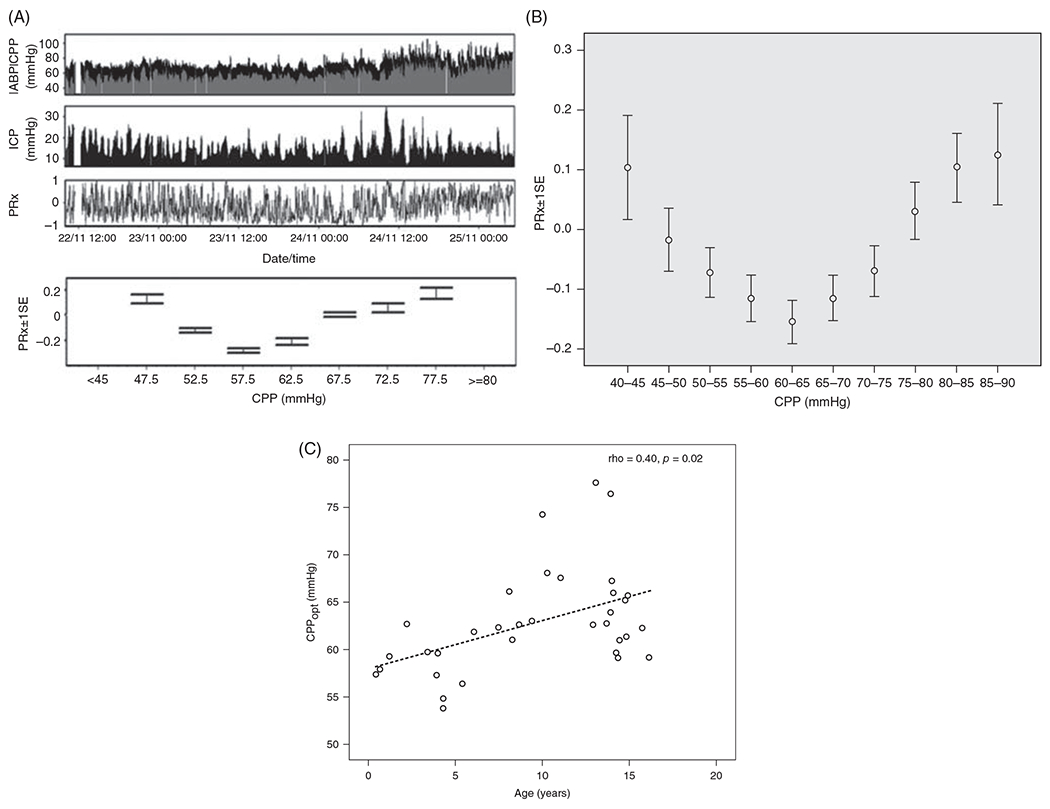
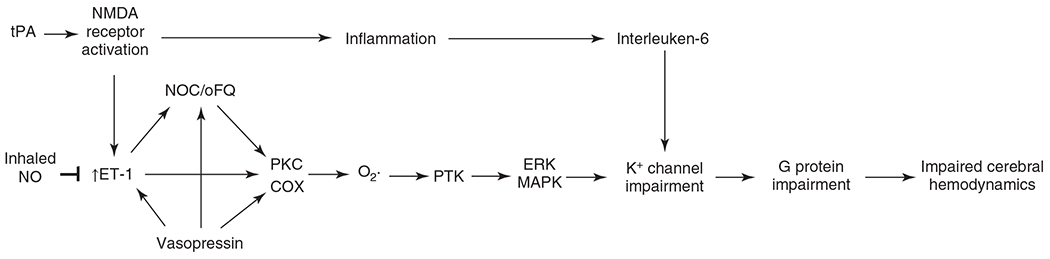
The cerebral microcirculation undergoes dynamic changes in parallel with the development of neurons, glia, and their energy metabolism throughout gestation and postnatally. Cerebral blood flow (CBF), oxygen consumption, and glucose consumption are as low as 20% of adult levels in humans born prematurely but eventually exceed adult levels at ages 3 to 11 years, which coincide with the period of continued brain growth, synapse formation, synapse pruning, and myelination. Neurovascular coupling to sensory activation is present but attenuated at birth. By 2 postnatal months, the increase in CBF often is disproportionately smaller than the increase in oxygen consumption, in contrast to the relative hyperemia seen in adults. Vascular smooth muscle myogenic tone increases in parallel with developmental increases in arterial pressure. CBF autoregulatory response to increased arterial pressure is intact at birth but has a more limited range with arterial hypotension. Hypoxia-induced vasodilation in preterm fetal sheep with low oxygen consumption does not sustain cerebral oxygen transport, but the response becomes better developed for sustaining oxygen transport by term. Nitric oxide tonically inhibits vasomotor tone, and glutamate receptor activation can evoke its release in lambs and piglets. In piglets, astrocyte-derived carbon monoxide plays a central role in vasodilation evoked by glutamate, ADP, and seizures, and prostanoids play a large role in endothelial-dependent and hypercapnic vasodilation. Overall, homeostatic mechanisms of CBF regulation in response to arterial pressure, neuronal activity, carbon dioxide, and oxygenation are present at birth but continue to develop postnatally as neurovascular signaling pathways are dynamically altered and integrated. © 2021 American Physiological Society. Compr Physiol 11:1-62, 2021.
脑微循环与神经元、神经胶质及其能量代谢的发育同步发生动态变化。早产儿出生时脑血流量(CBF)、氧消耗和葡萄糖消耗仅为成人的 20%,但在 3 至 11 岁时最终超过成人水平,这与大脑持续生长、突触形成、突触修剪和髓鞘形成的时期相吻合。出生时,感觉激活的神经血管耦联存在但减弱。到 2 个月大时,CBF 的增加通常不成比例地小于氧消耗的增加,与成人中所见的相对充血形成对比。血管平滑肌肌源性张力与动脉压的发育性增加平行增加。出生时 CBF 的自动调节反应对动脉压升高是完整的,但在动脉低血压时范围更有限。低氧诱导的低氧消耗的早产胎儿羊的血管扩张不能维持脑氧输送,但到足月时,对维持氧输送的反应变得更加发达。一氧化氮持续抑制血管舒缩张力,谷氨酸受体激活可在羔羊和仔猪中诱发其释放。在仔猪中,星形胶质细胞衍生的一氧化碳在谷氨酸、ADP 和癫痫发作引起的血管扩张中起核心作用,前列腺素在内皮依赖性和高碳酸血症性血管扩张中起重要作用。总的来说,出生时就存在 CBF 调节对动脉压、神经元活动、二氧化碳和氧合作用的稳态机制,但随着神经血管信号通路的动态改变和整合,它们在出生后继续发育。© 2021 美国生理学会。综合生理学 11:1-62,2021。