Sir John Walsh Research Institute, University of Otagogrid.29980.3a, Dunedin, New Zealand.
Department of Food Science and Human Nutrition, Faculty of Agriculture, Ryukoku University, Shiga, Japan.
Antimicrob Agents Chemother. 2022 Jan 18;66(1):e0174821. doi: 10.1128/AAC.01748-21. Epub 2021 Nov 15.
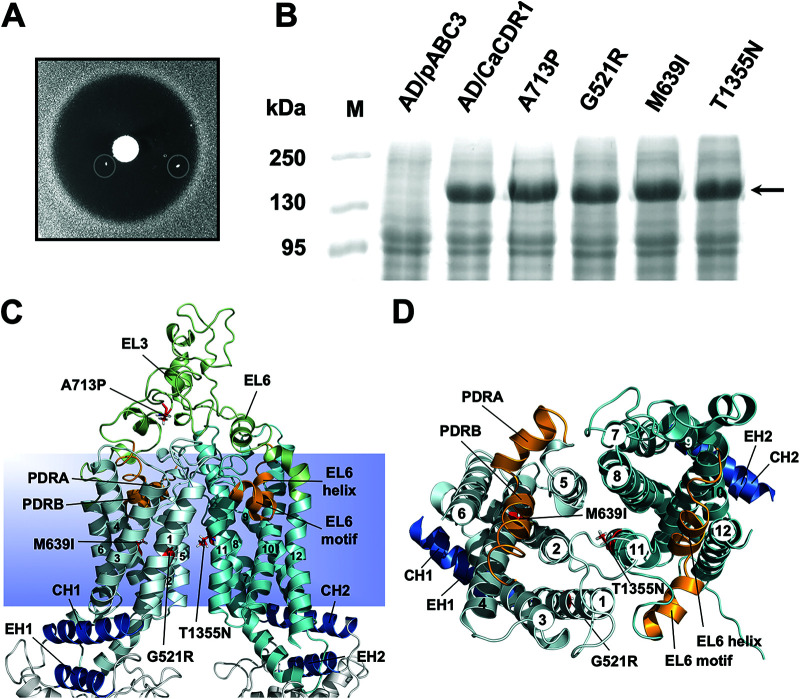
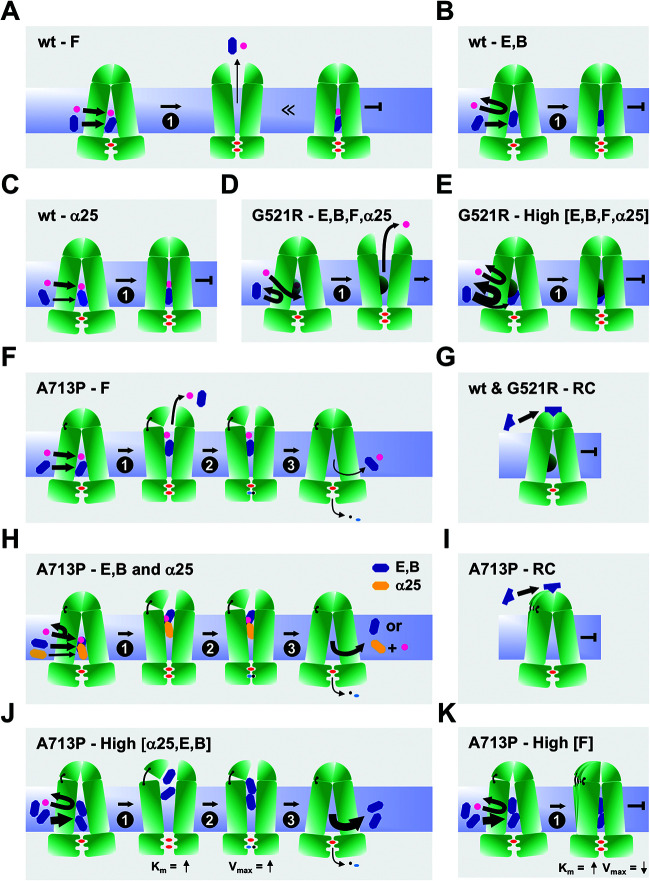
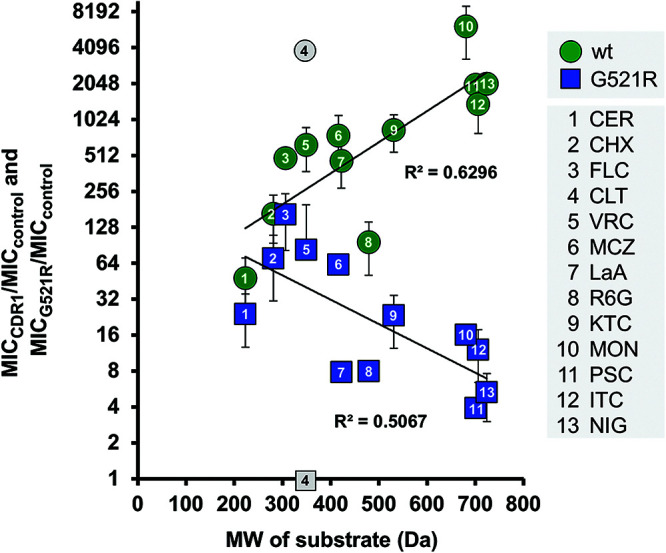
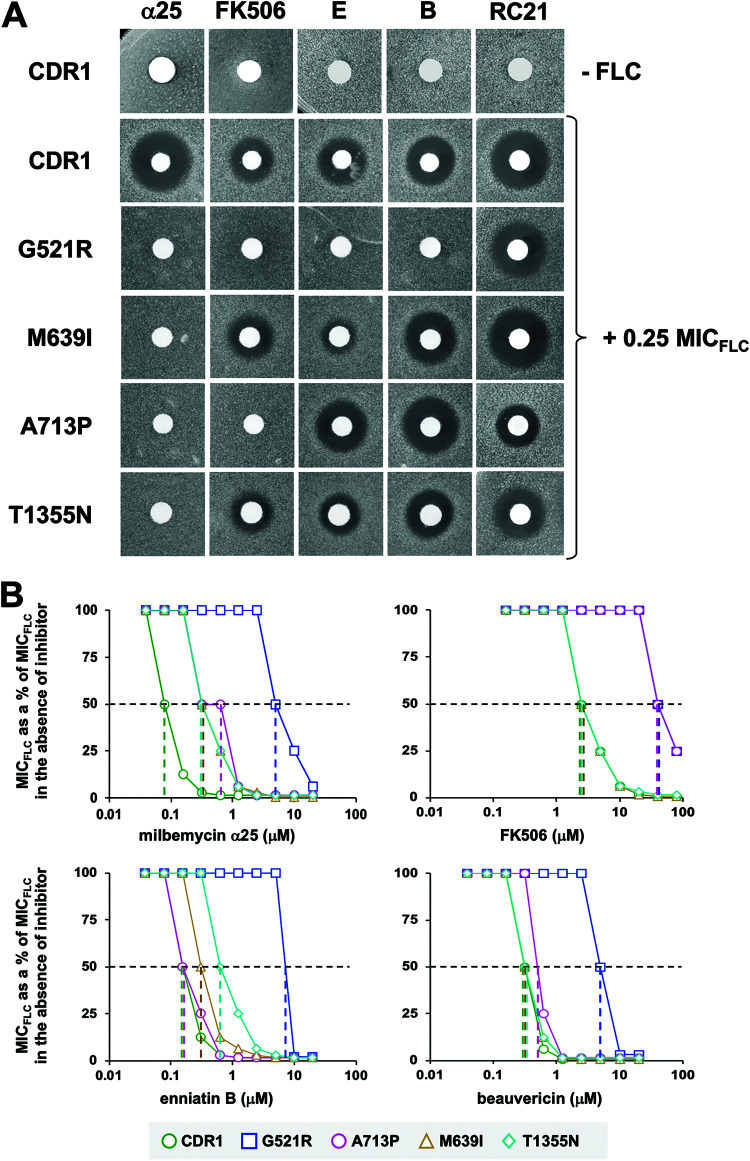
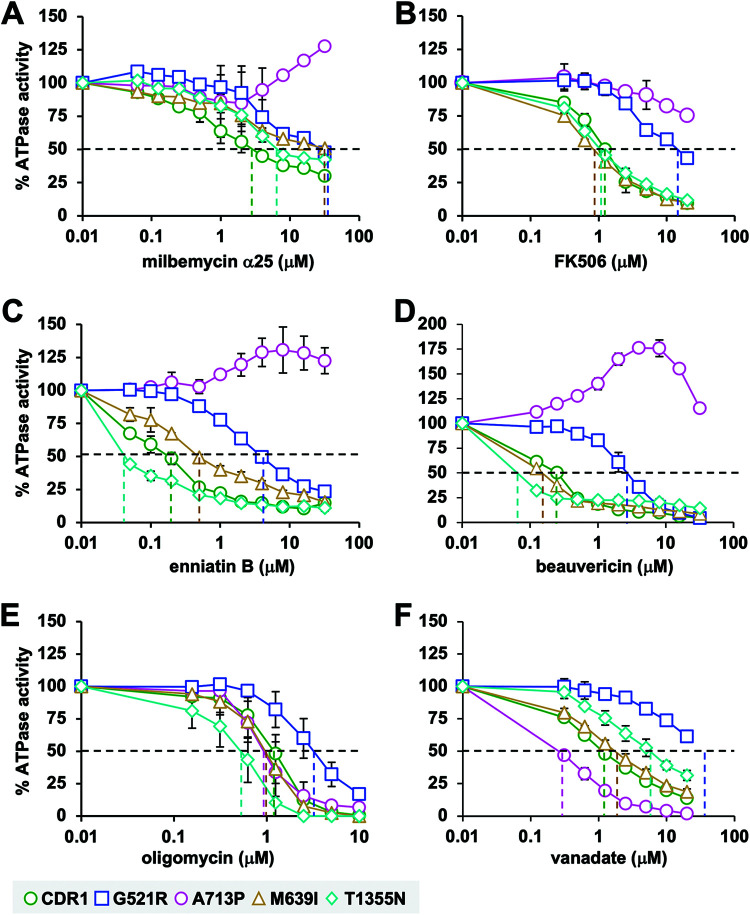
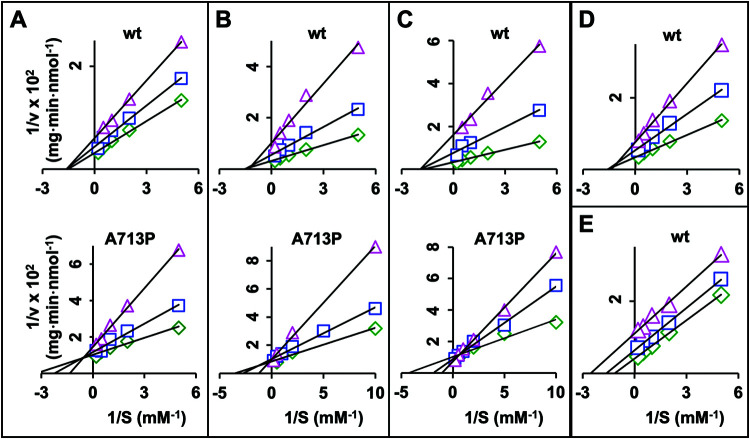
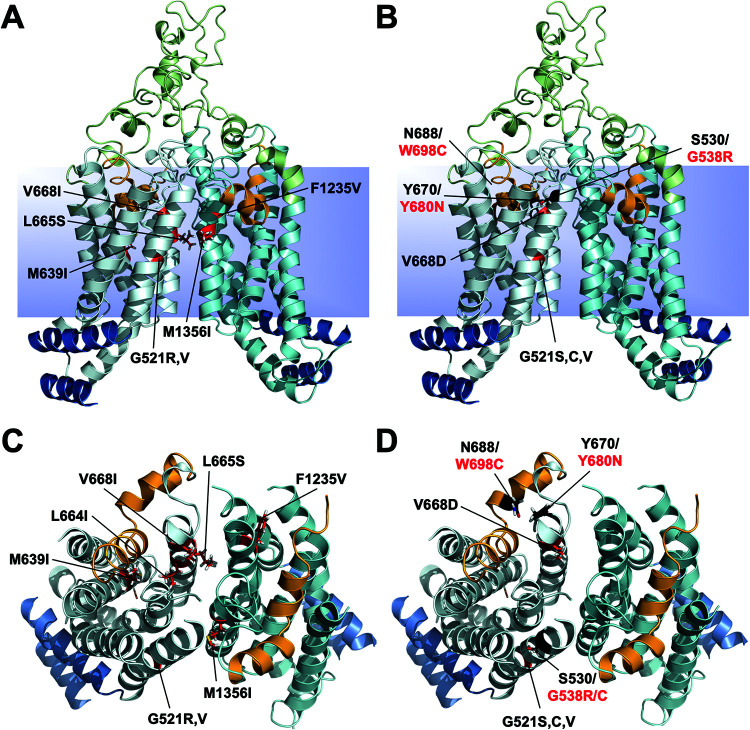
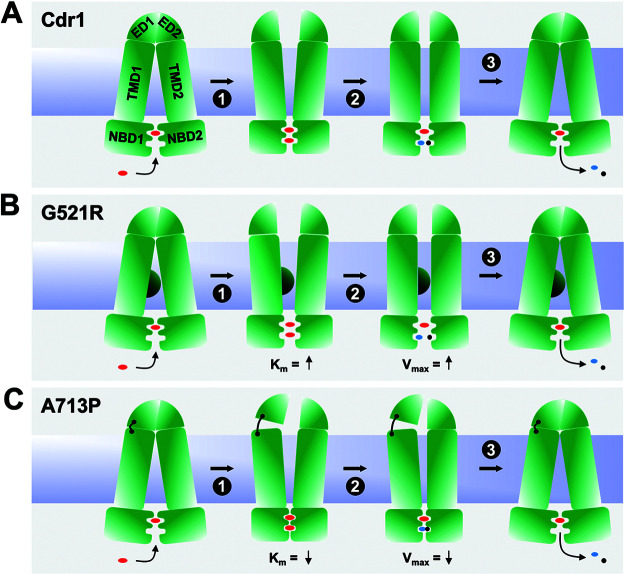
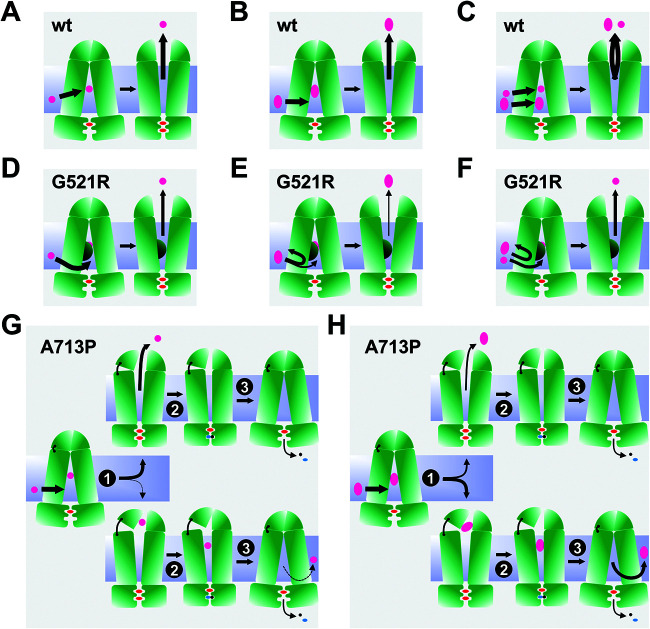
Overexpression of ATP-binding cassette (ABC) transporters is a major cause of drug resistance in fungal pathogens. Milbemycins, enniatin B, beauvericin, and FK506 are promising leads for broad-spectrum fungal multidrug efflux pump inhibitors. The characterization of naturally generated inhibitor-resistant mutants is a powerful tool to elucidate structure-activity relationships in ABC transporters. We isolated 20 Saccharomyces cerevisiae mutants overexpressing Candida albicans ABC pump Cdr1 variants resistant to fluconazole efflux inhibition by milbemycin α25 (8 mutants), enniatin B (8), or beauvericin (4). The 20 mutations were in just 9 residues at the centers of transmembrane segment 1 (TMS1) (6 mutations), TMS4 (4), TMS5 (4), TMS8 (1), and TMS11 (2) and in A713P (3), a previously reported FK506-resistant "hot spot 1" mutation in extracellular loop 3. Six Cdr1-G521S/C/V/R (TMS1) variants were resistant to all four inhibitors, four Cdr1-M639I (TMS4) variants were resistant to milbemycin α25 and enniatin B, and two Cdr1-V668I/D (TMS5) variants were resistant to enniatin B and beauvericin. The eight milbemycin α25-resistant mutants were altered in four amino acids as follows: G521R, M639I, A713P, and T1355N (TMS11). These four Cdr1 variants responded differently to various types of inhibitors, and each exhibited altered substrate specificity and kinetic properties. The data infer an entry gate function for Cdr1-G521 and a role for Cdr1-A713 in the constitutively high Cdr1 ATPase activity. Cdr1-M639I and -T1355N possibly cause inhibitor resistance by altering TMS contacts near the substrate/inhibitor-binding pocket. Models for the interactions of substrates and different types of inhibitors with Cdr1 at various stages of the transport cycle are presented.
多药外排泵的过度表达是真菌病原体产生耐药性的主要原因。米尔贝肟、恩镰菌素 B、伏马菌素和 FK506 是广谱抗真菌多药外排泵抑制剂的有希望的先导化合物。天然产生的抑制剂抗性突变体的特征描述是阐明 ABC 转运蛋白结构-活性关系的有力工具。我们分离了 20 个过度表达白念珠菌 ABC 泵 Cdr1 变体的酿酒酵母突变体,这些变体对米贝肟 α25(8 个突变体)、恩镰菌素 B(8 个)或伏马菌素(4 个)的氟康唑外排抑制作用具有抗性。这 20 个突变仅发生在跨膜片段 1(TMS1)(6 个突变)、TMS4(4 个)、TMS5(4 个)、TMS8(1 个)和 TMS11(2 个)的中心以及先前报道的细胞外环 3 中的 FK506 抗性“热点 1”突变 A713P。6 个 Cdr1-G521S/C/V/R(TMS1)变体对所有四种抑制剂均具有抗性,4 个 Cdr1-M639I(TMS4)变体对米贝肟 α25 和恩镰菌素 B 具有抗性,2 个 Cdr1-V668I/D(TMS5)变体对恩镰菌素 B 和伏马菌素具有抗性。8 个米贝肟 α25 抗性突变体在以下四个氨基酸中发生改变:G521R、M639I、A713P 和 T1355N(TMS11)。这四个 Cdr1 变体对不同类型的抑制剂有不同的反应,并且每个变体都表现出改变的底物特异性和动力学特性。数据推断 Cdr1-G521 具有入口门功能,而 Cdr1-A713 在 Cdr1 ATP 酶的固有高活性中起作用。Cdr1-M639I 和 -T1355N 可能通过改变靠近底物/抑制剂结合口袋的 TMS 接触来引起抑制剂抗性。提出了在运输循环的各个阶段,底物和不同类型抑制剂与 Cdr1 相互作用的模型。