Department of Structural Biochemistry, Max Planck Institute of Molecular Physiology, Dortmund, Germany.
Department of Pharmacology and Therapeutics and the Myology Institute, University of Florida, Gainesville, United States.
Elife. 2021 Nov 23;10:e73724. doi: 10.7554/eLife.73724.
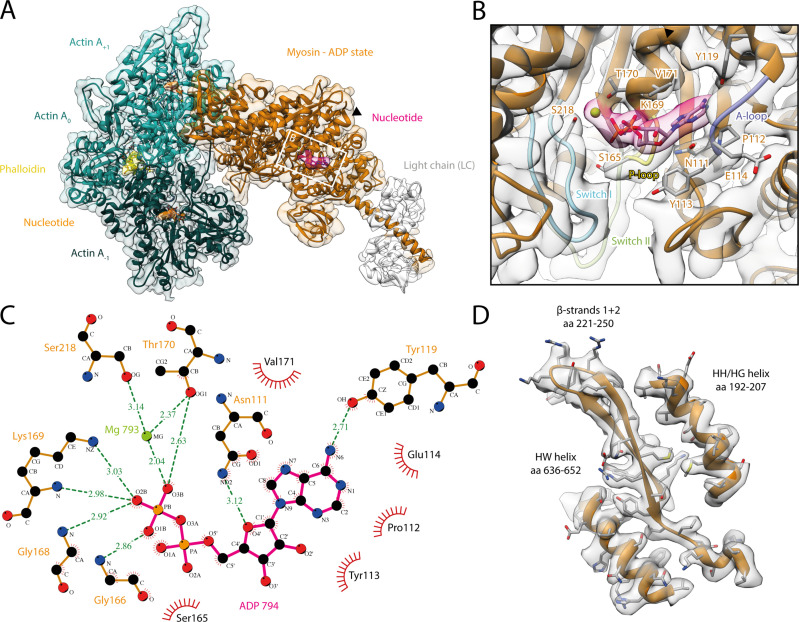
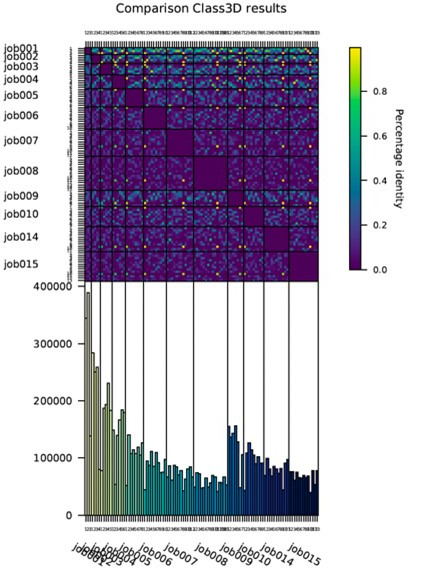
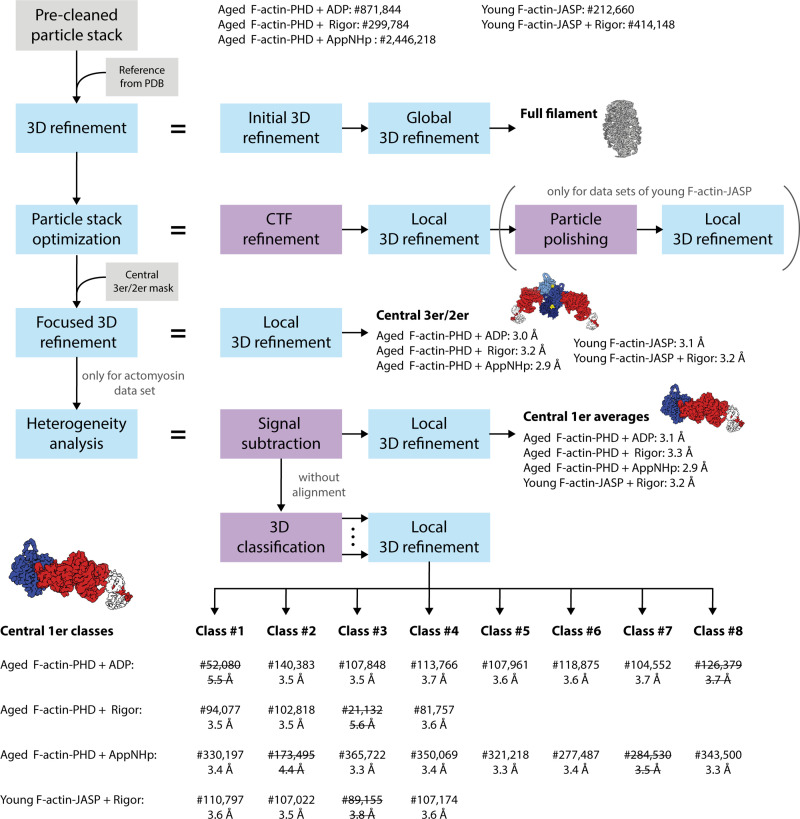
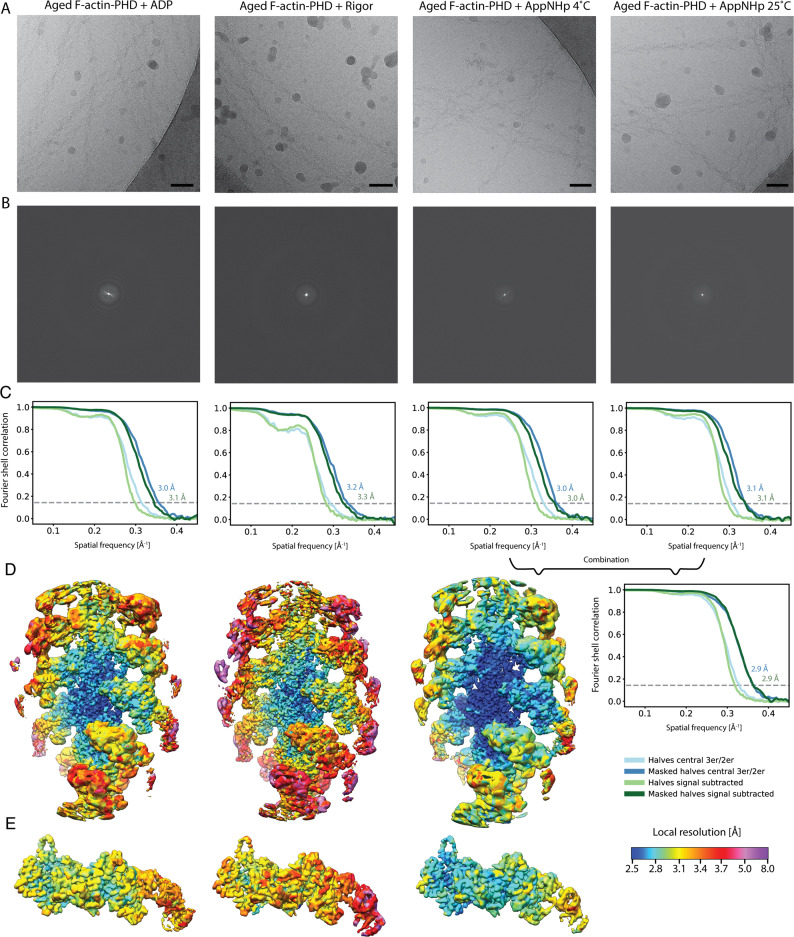
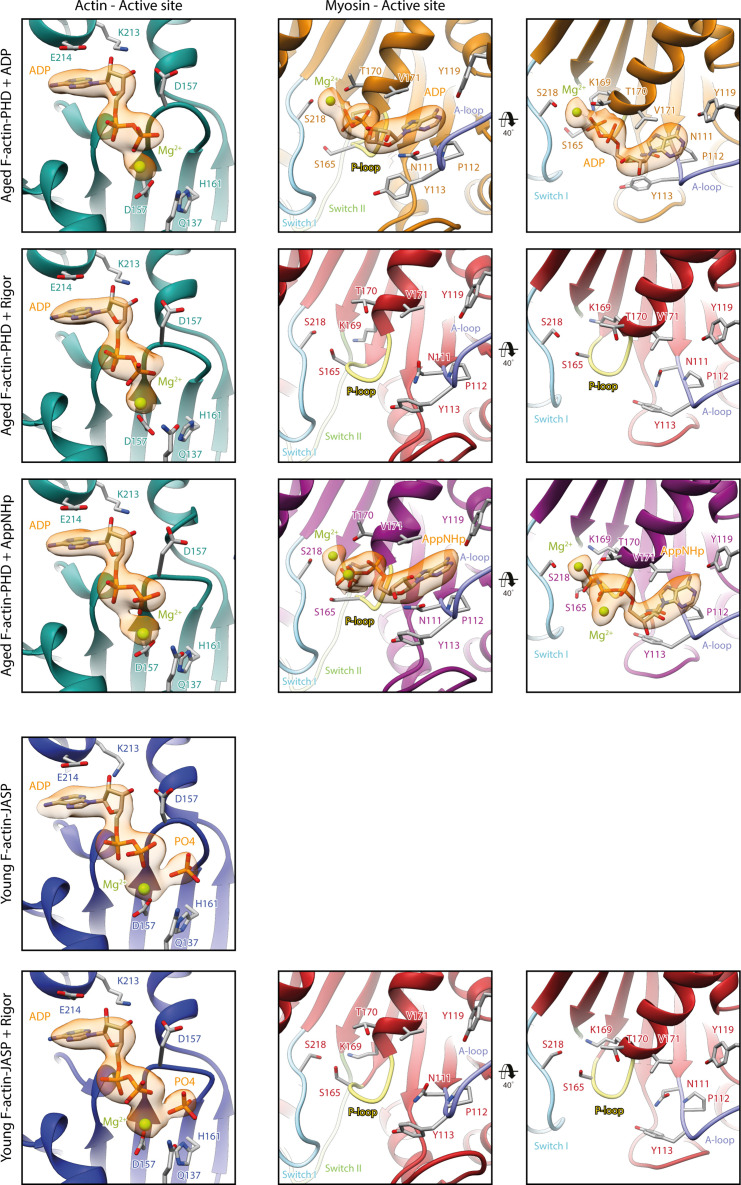
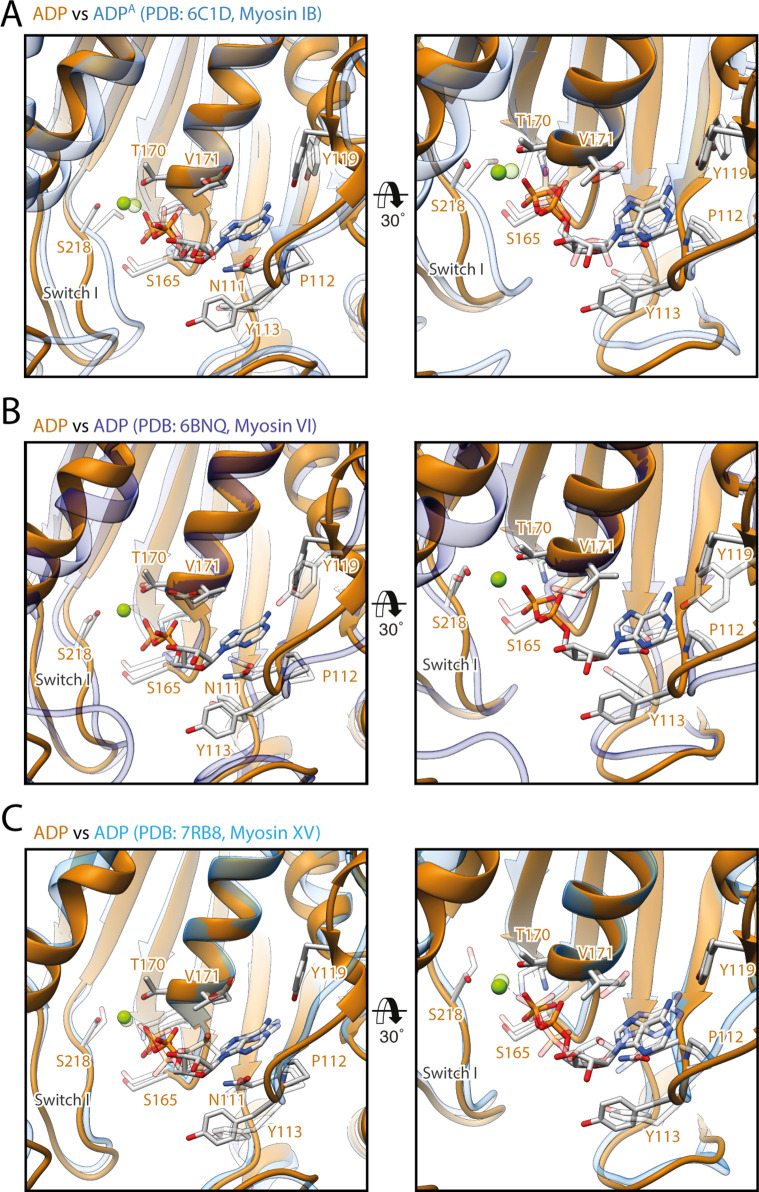
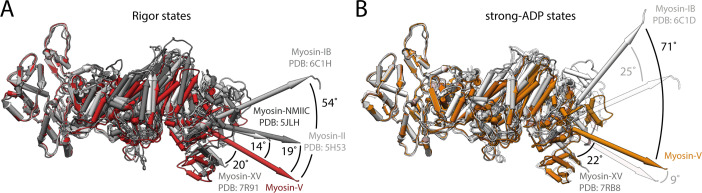
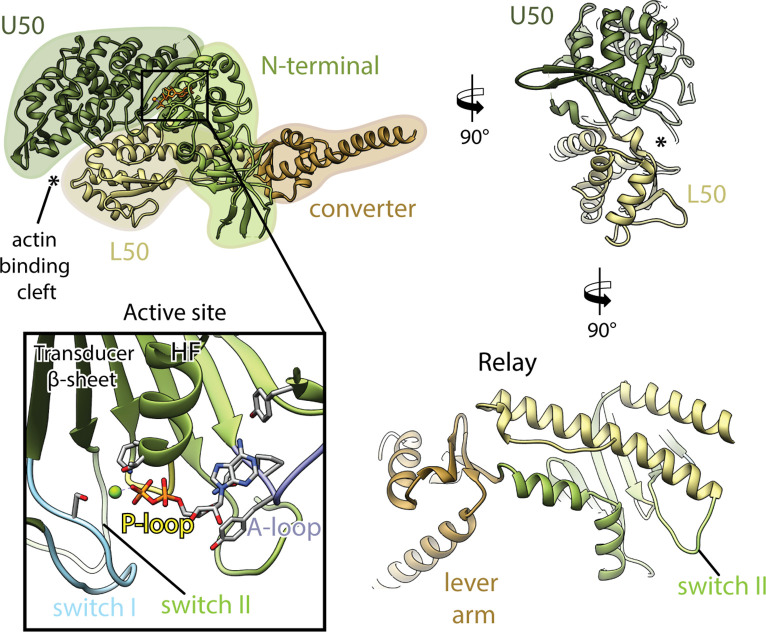
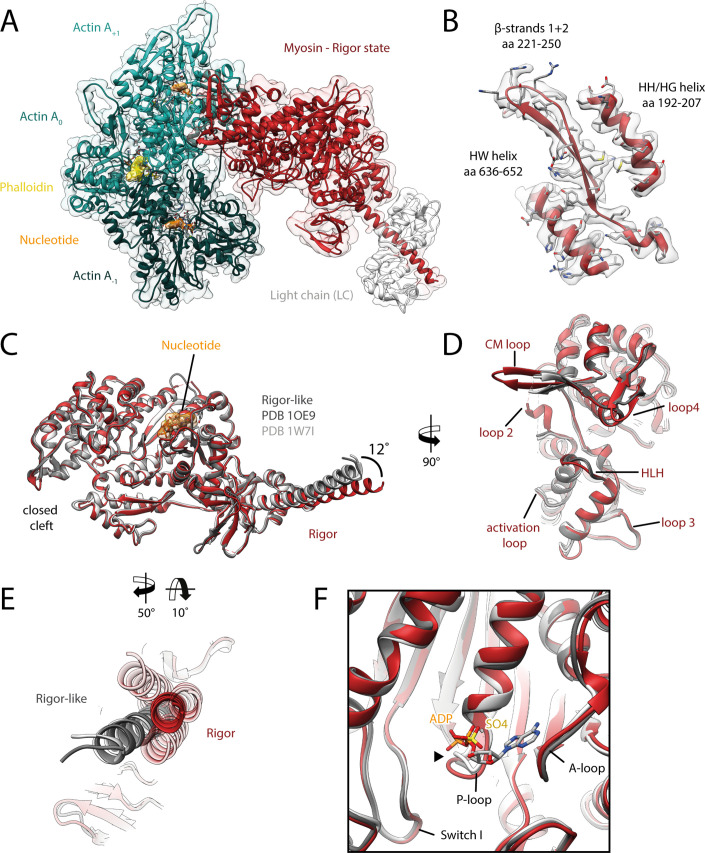
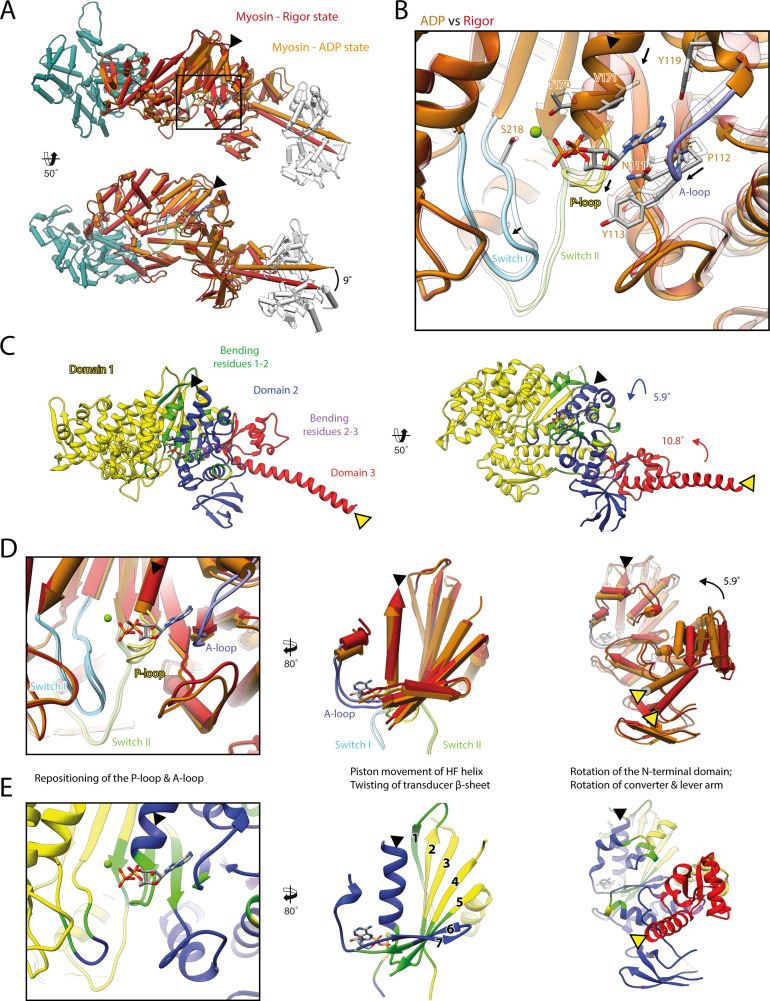
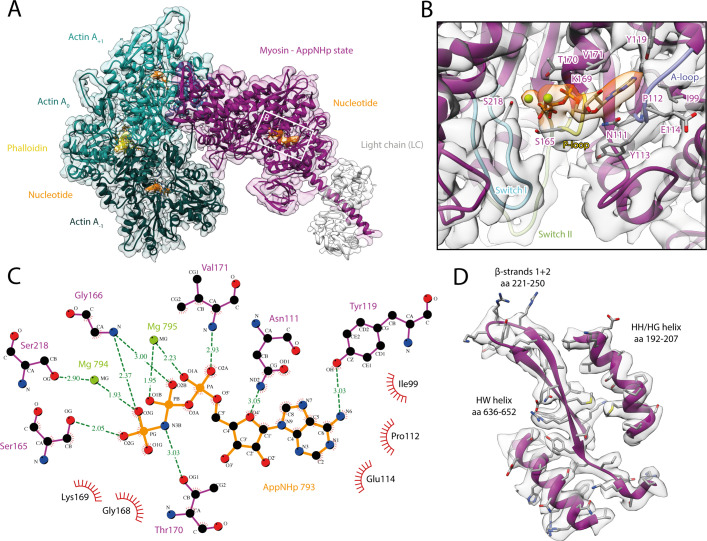
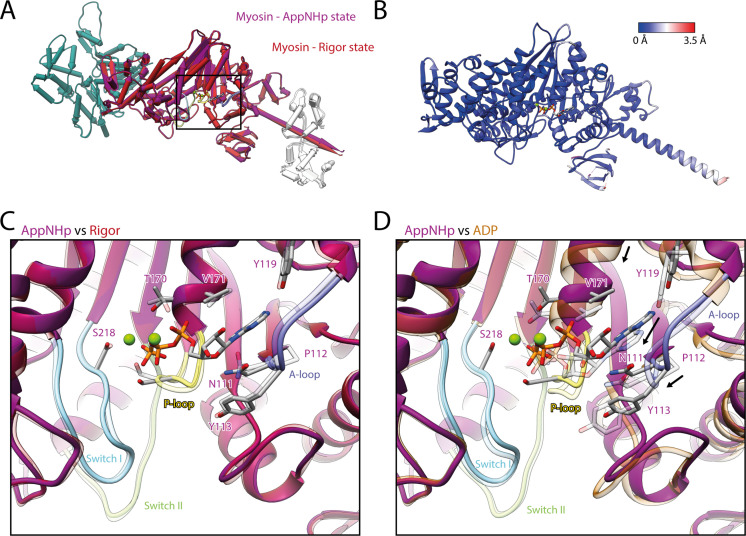
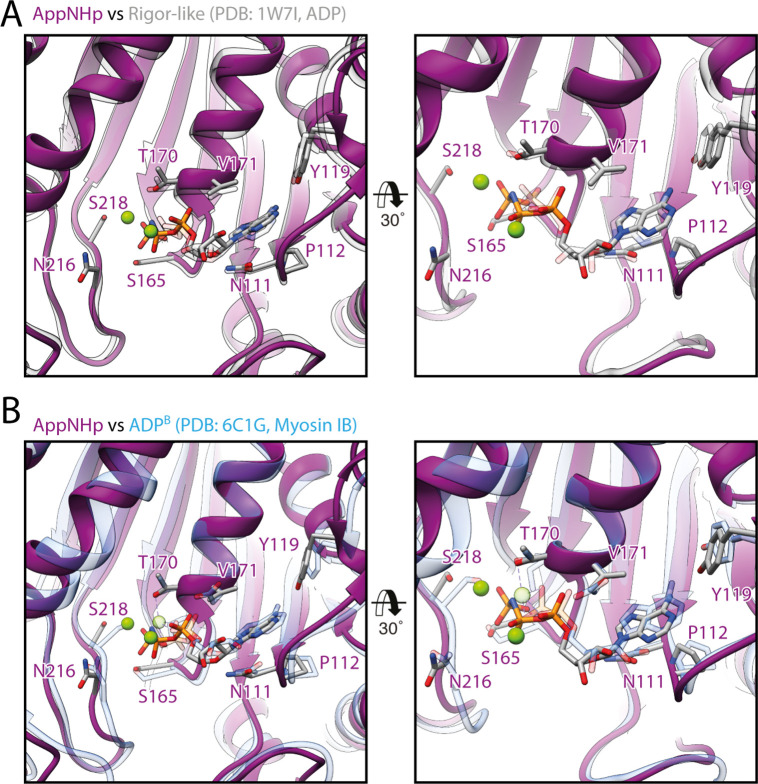
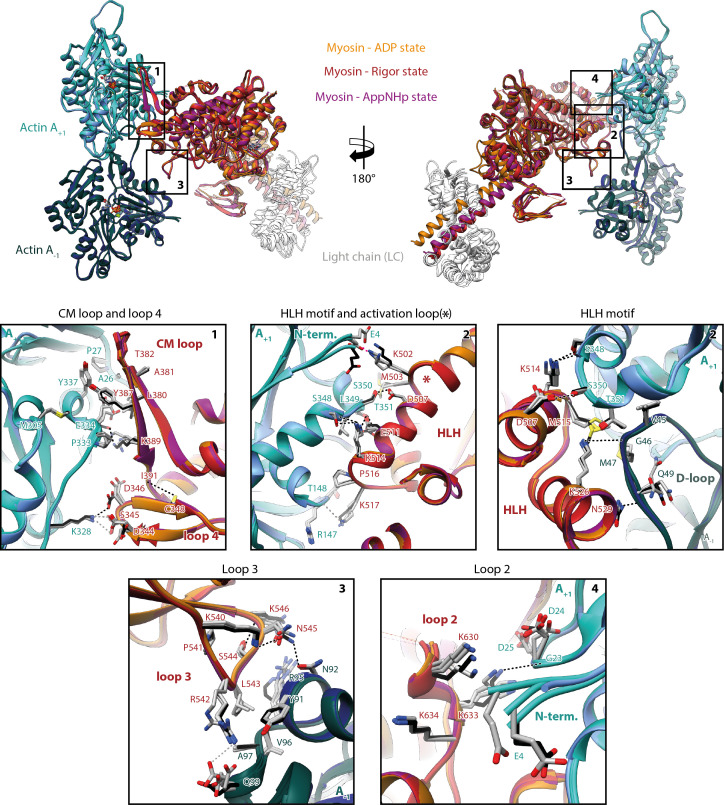
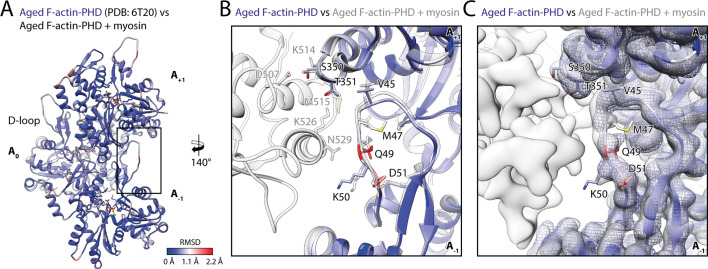
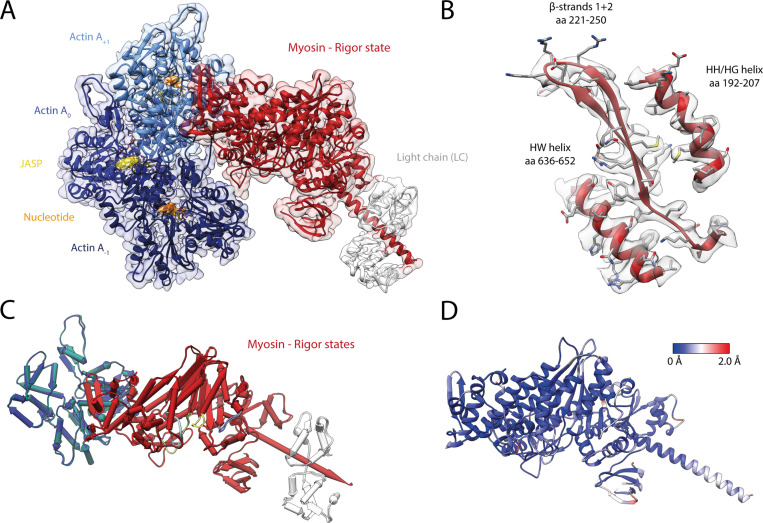
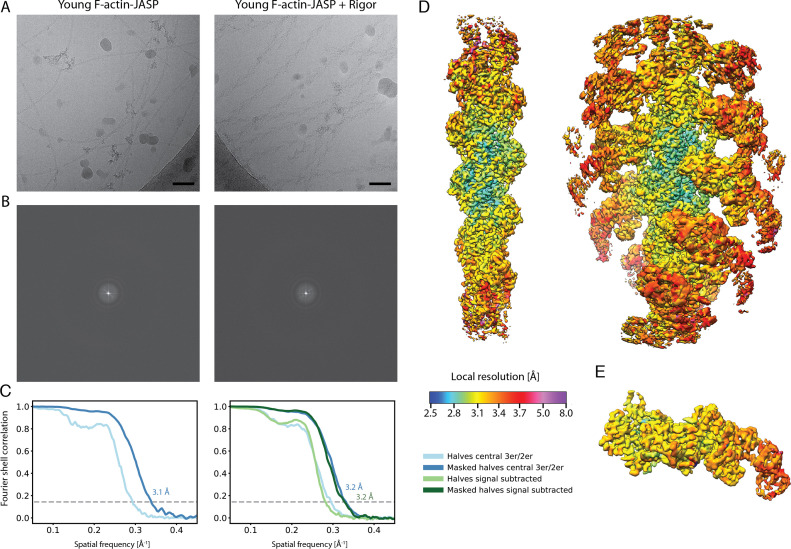
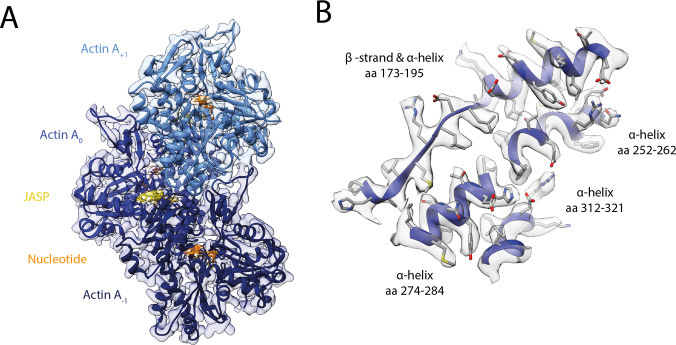
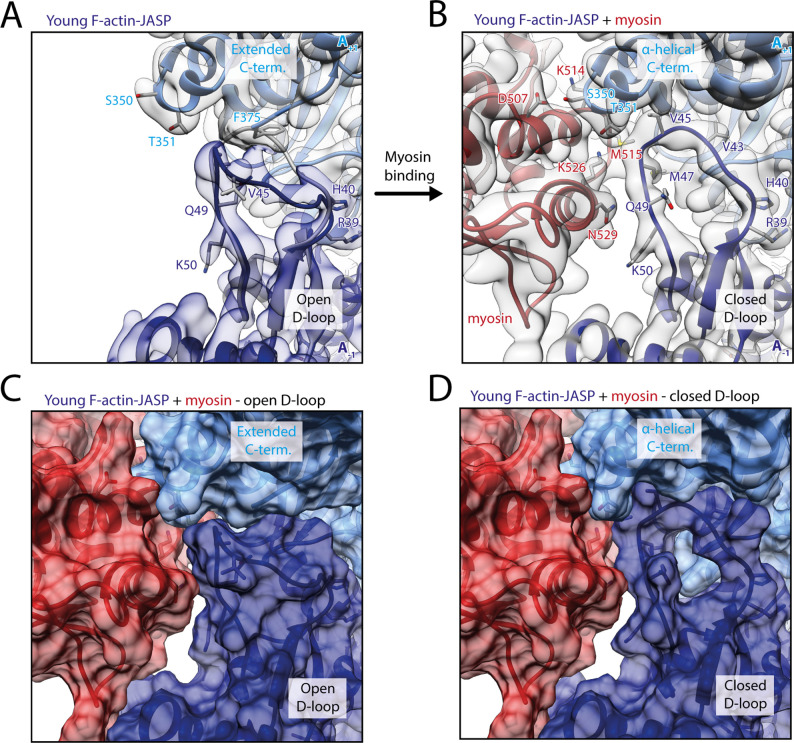
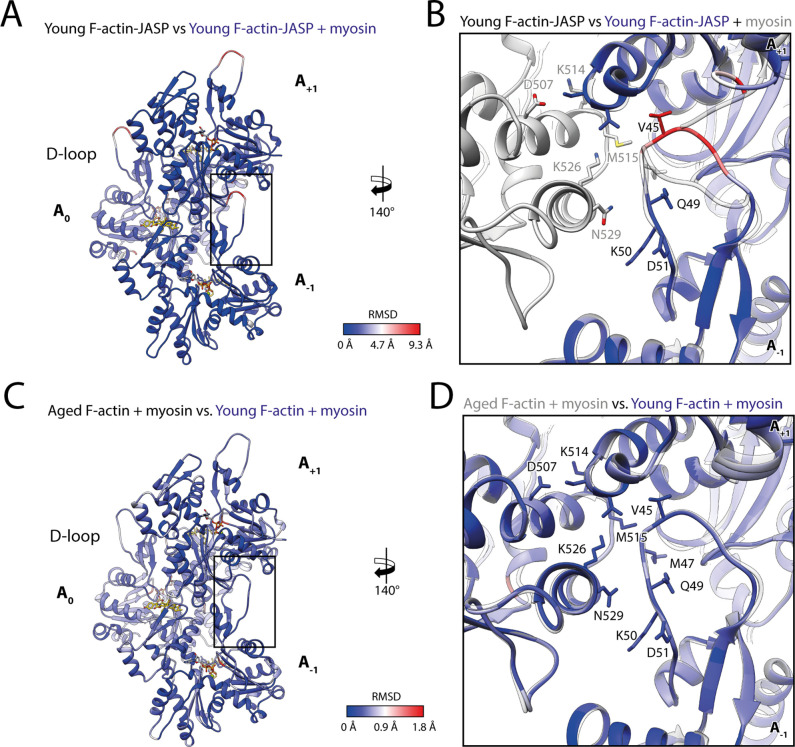
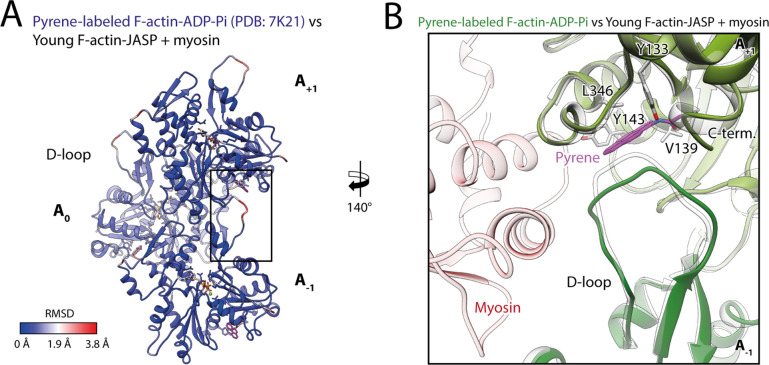
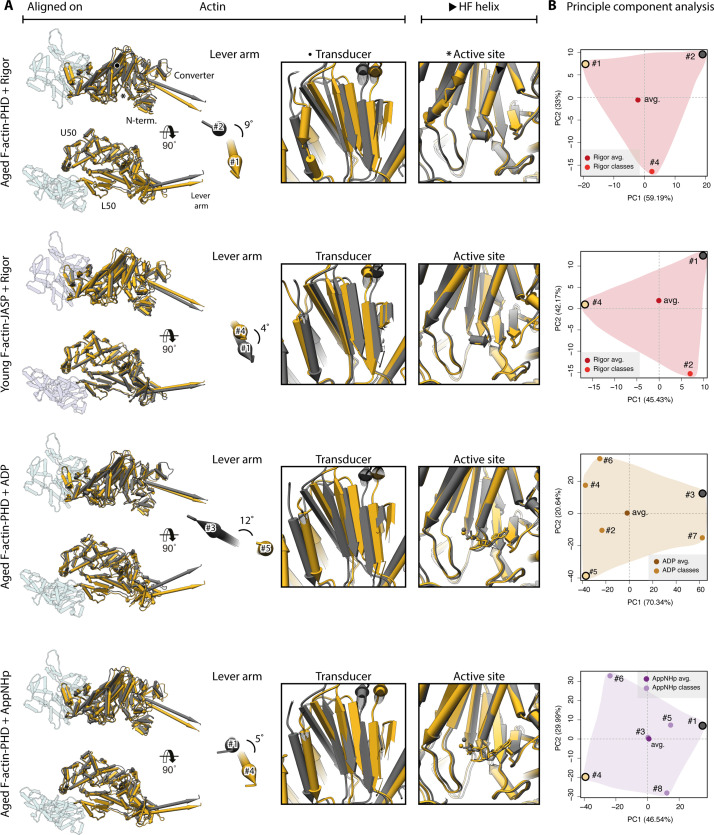
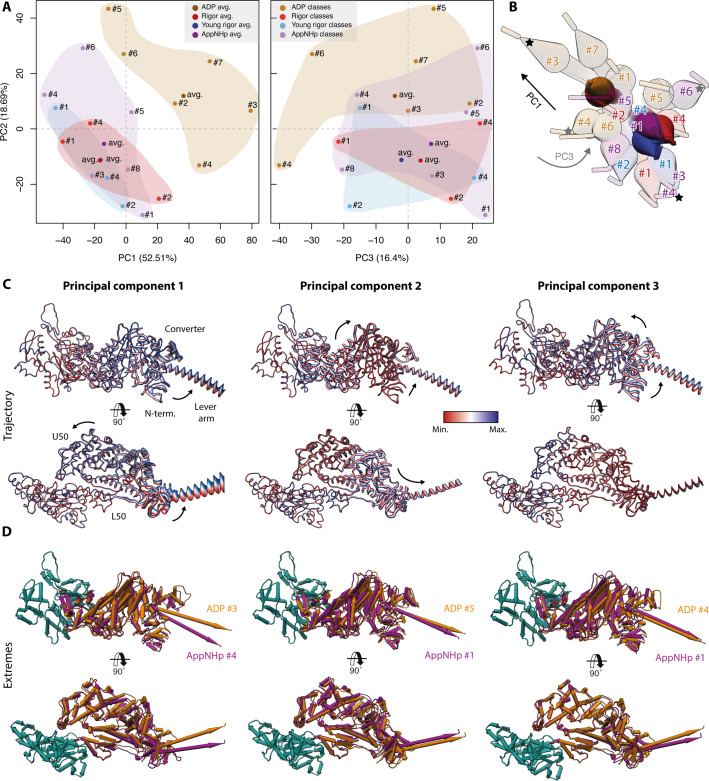
The molecular motor myosin undergoes a series of major structural transitions during its force-producing motor cycle. The underlying mechanism and its coupling to ATP hydrolysis and actin binding are only partially understood, mostly due to sparse structural data on actin-bound states of myosin. Here, we report 26 high-resolution cryo-EM structures of the actomyosin-V complex in the strong-ADP, rigor, and a previously unseen post-rigor transition state that binds the ATP analog AppNHp. The structures reveal a high flexibility of myosin in each state and provide valuable insights into the structural transitions of myosin-V upon ADP release and binding of AppNHp, as well as the actomyosin interface. In addition, they show how myosin is able to specifically alter the structure of F-actin.
分子马达肌球蛋白在产生力的运动循环中经历一系列主要的结构转变。其潜在机制及其与 ATP 水解和肌动蛋白结合的偶联在很大程度上尚不清楚,主要是因为关于肌球蛋白结合肌动蛋白状态的结构数据稀疏。在这里,我们报告了肌球蛋白 - 肌动球蛋白 V 复合物在强 ADP、僵硬和以前未见的后僵硬过渡状态下的 26 个高分辨率冷冻电镜结构,该复合物结合了 ATP 类似物 AppNHp。这些结构揭示了每种状态下肌球蛋白的高灵活性,并为肌球蛋白 V 在 ADP 释放和 AppNHp 结合以及肌球蛋白 - 肌动蛋白界面时的结构转变提供了有价值的见解。此外,它们还展示了肌球蛋白如何能够特异性地改变 F- 肌动蛋白的结构。