Department of Cellular and Molecular Medicine, San Diego, United States.
Department of Biochemistry and Molecular Genetics, University of Colorado Denver, Aurora, United States.
Elife. 2021 Dec 9;10:e74538. doi: 10.7554/eLife.74538.
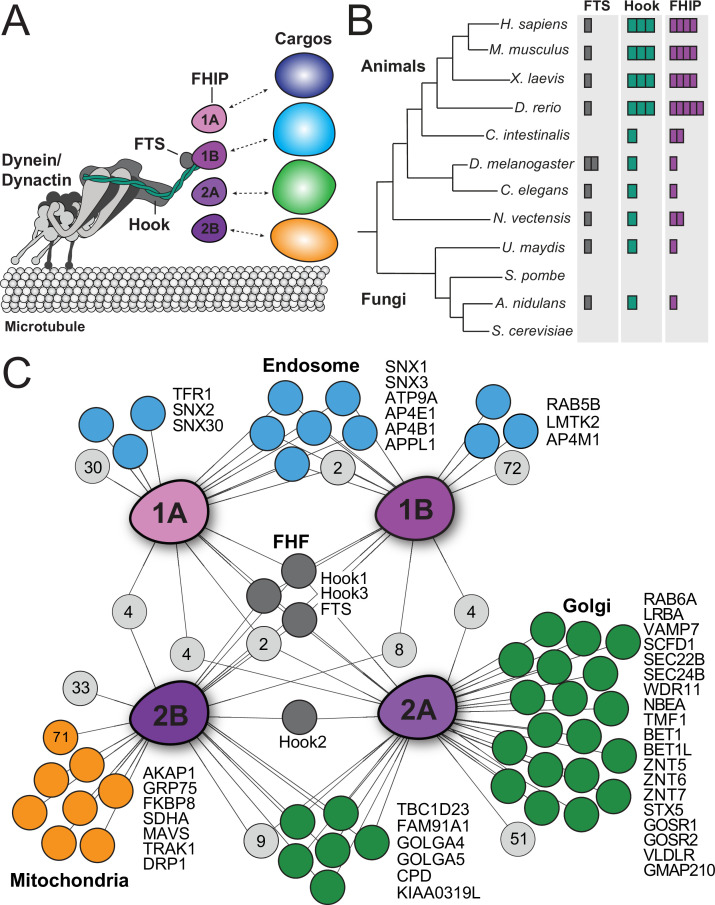
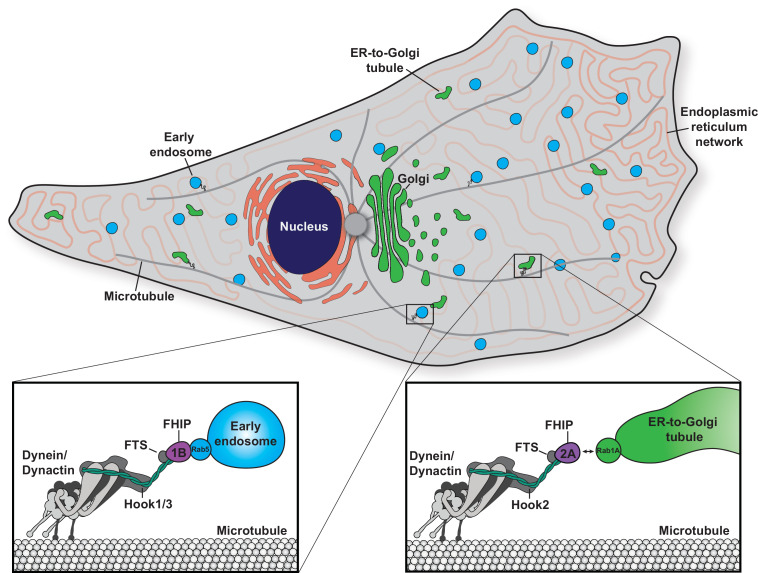
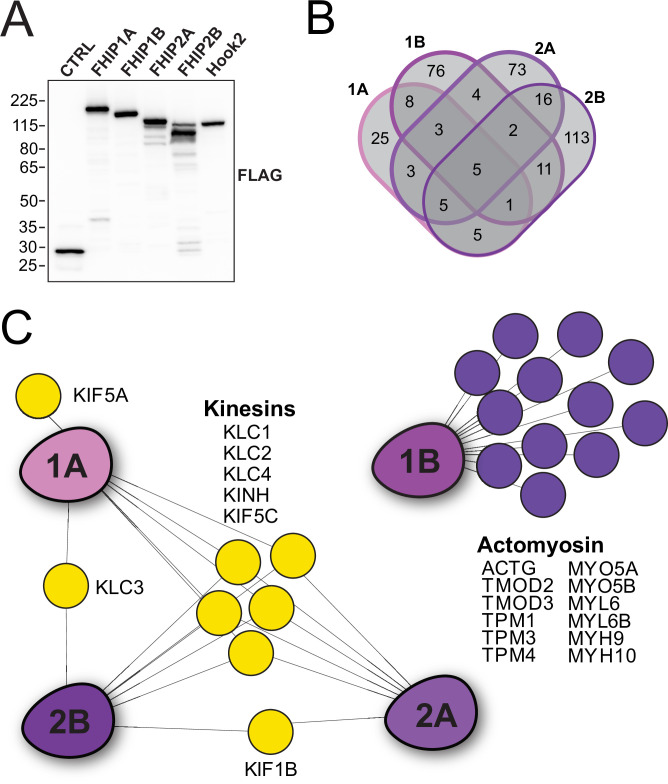
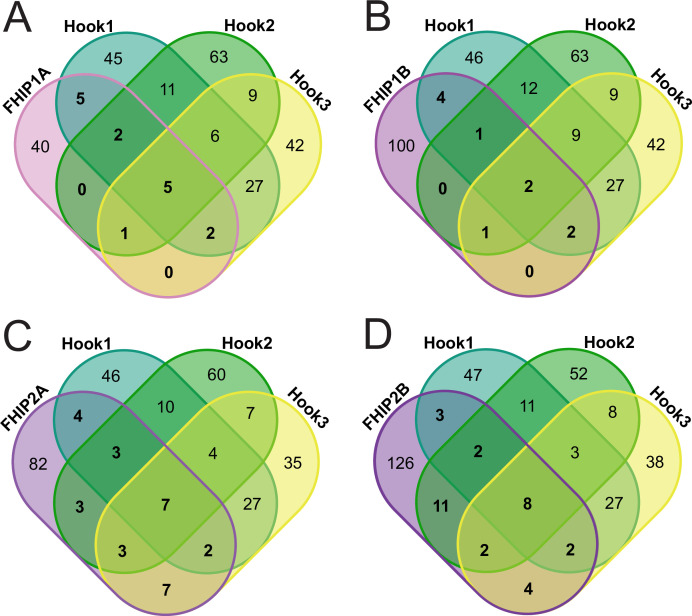
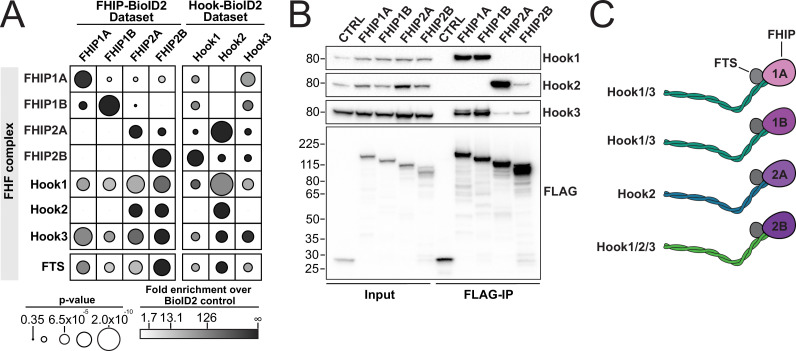
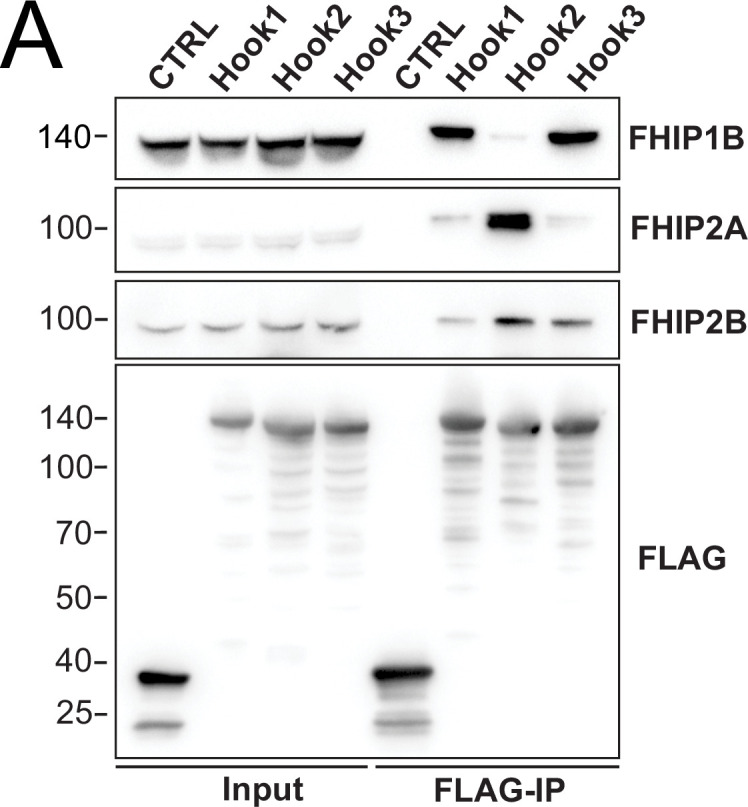
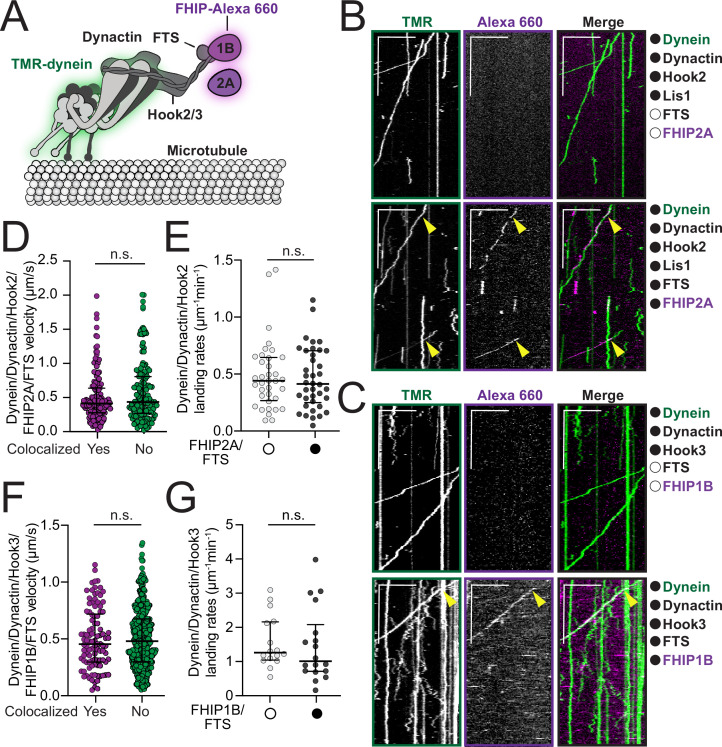
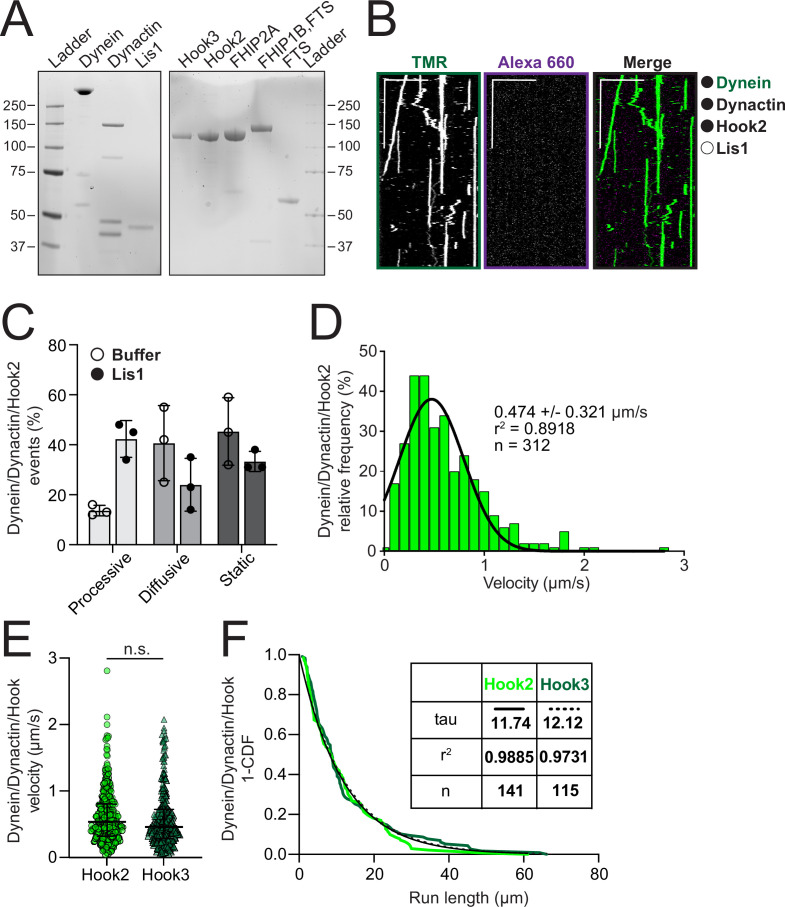
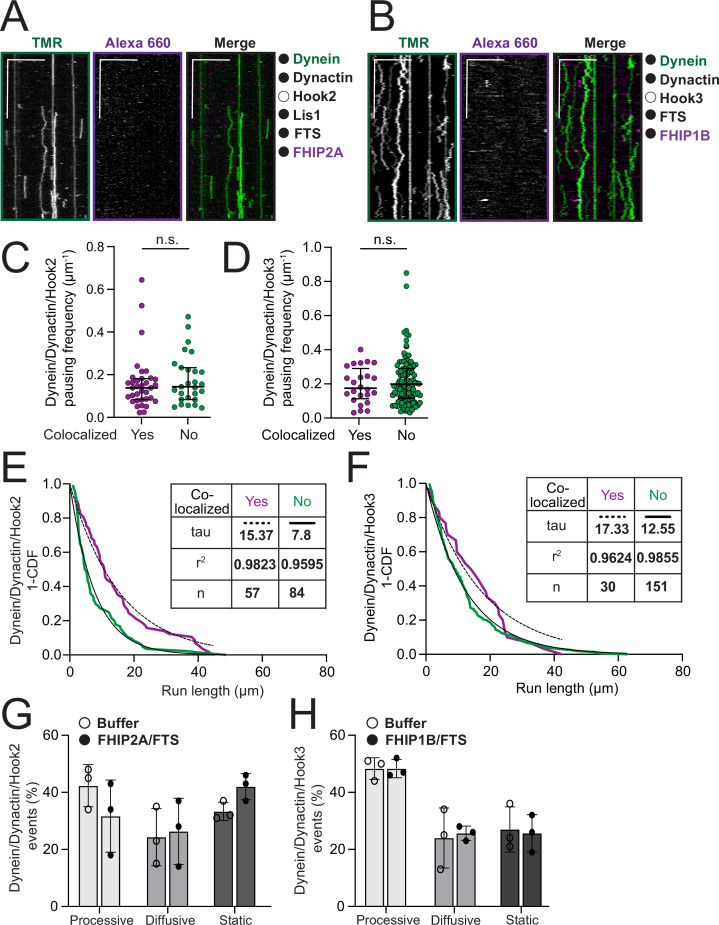
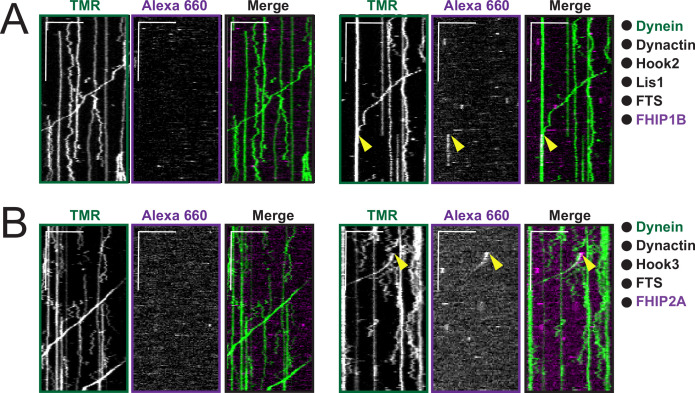
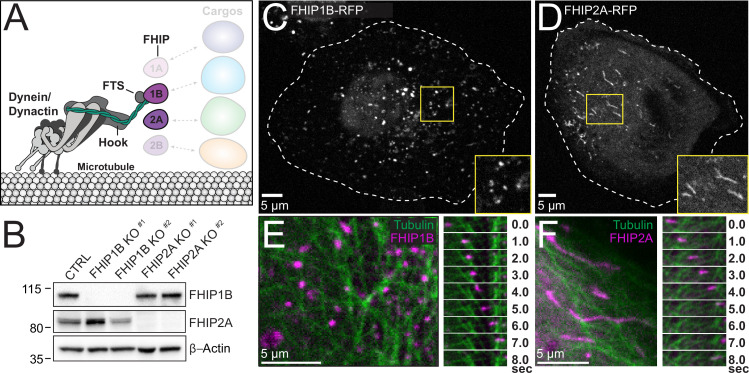
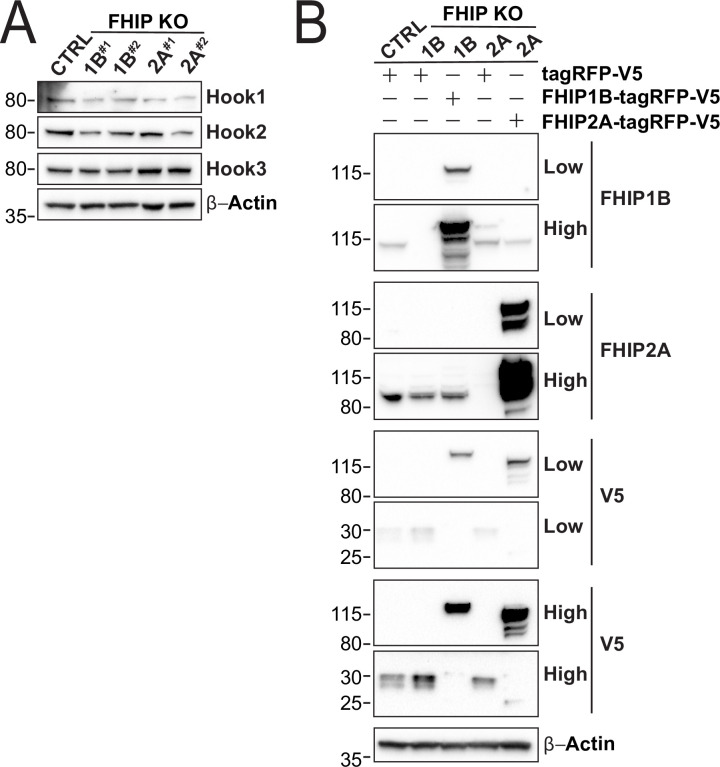
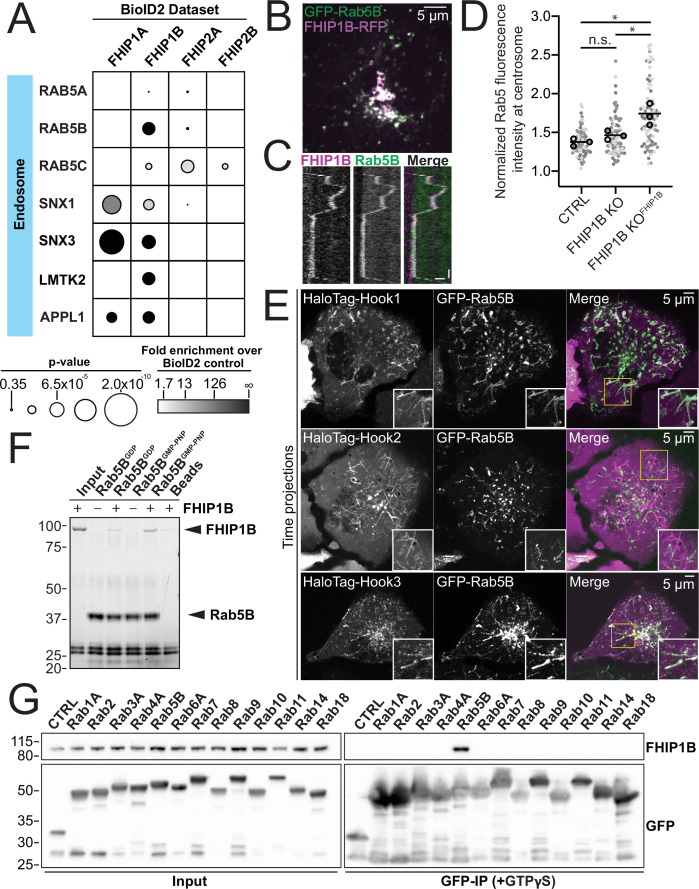
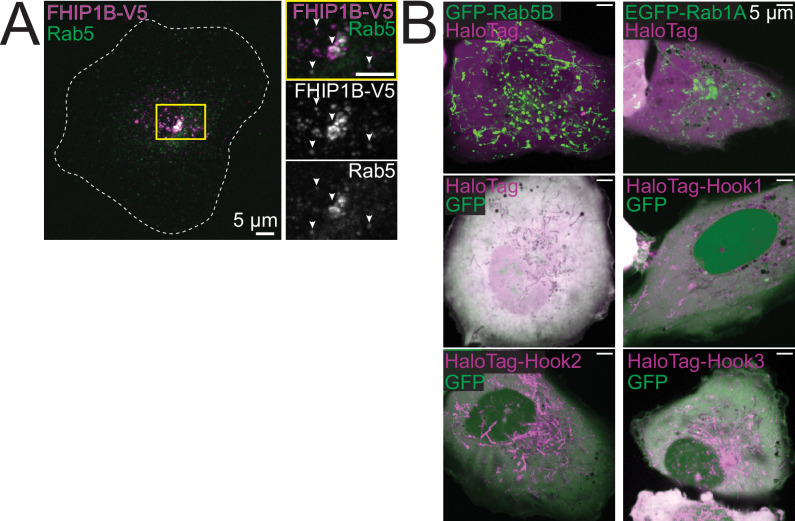
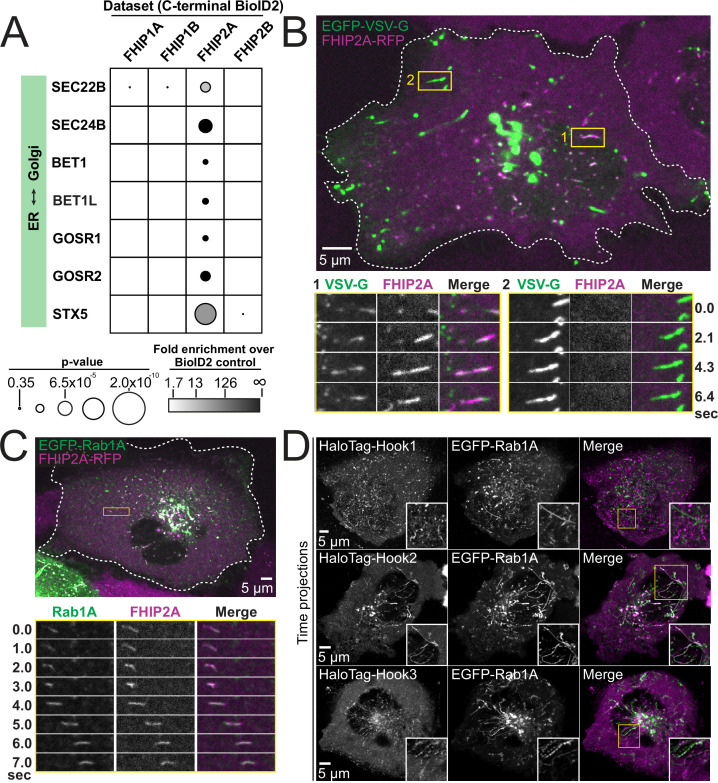
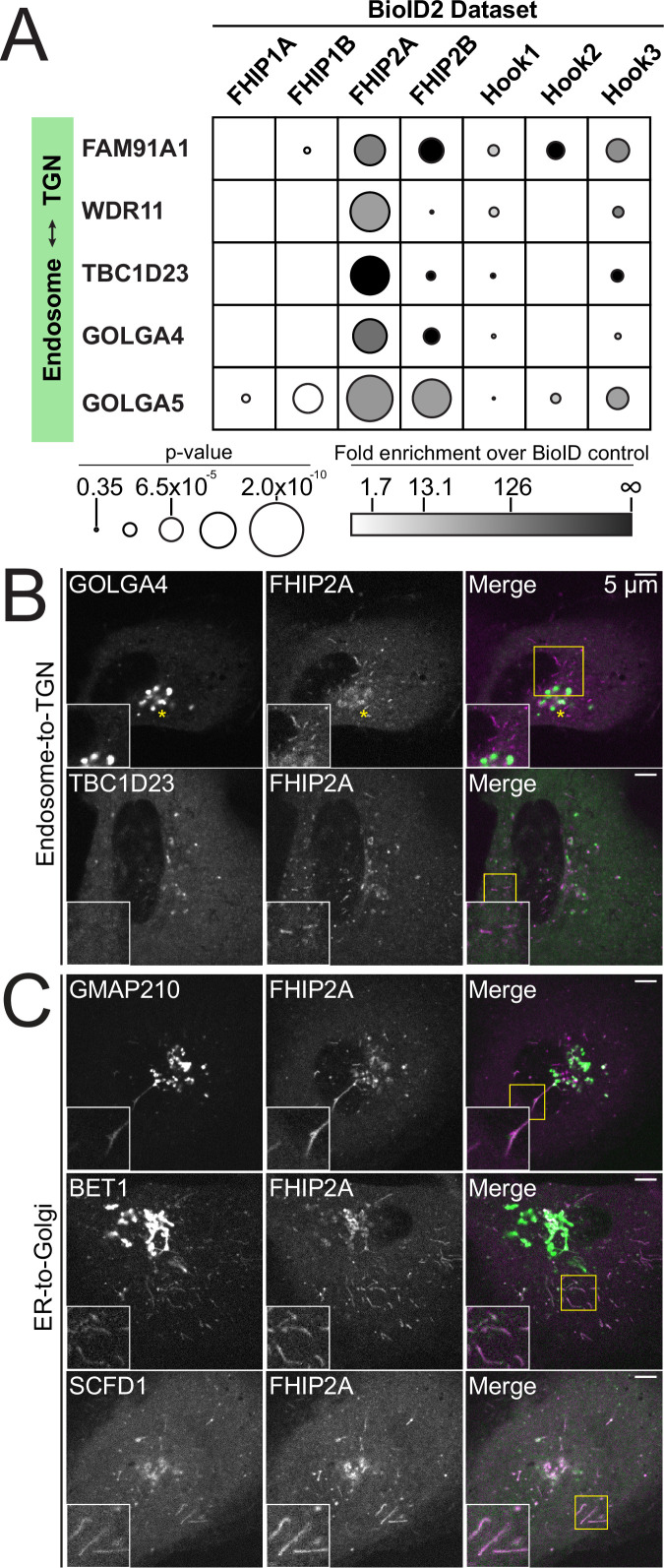
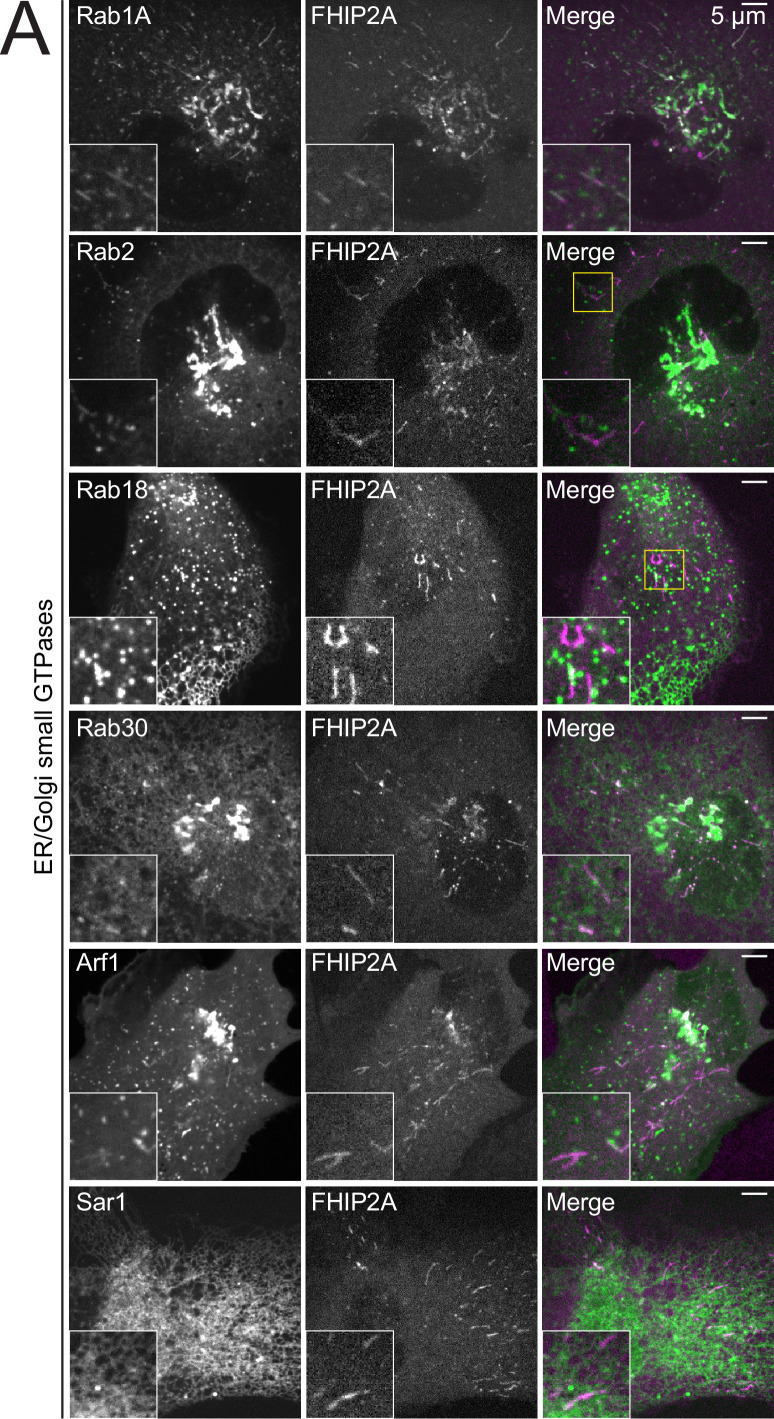
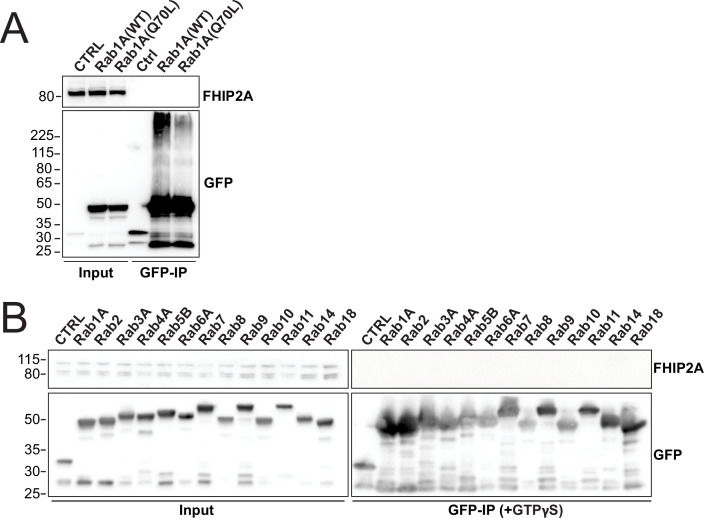
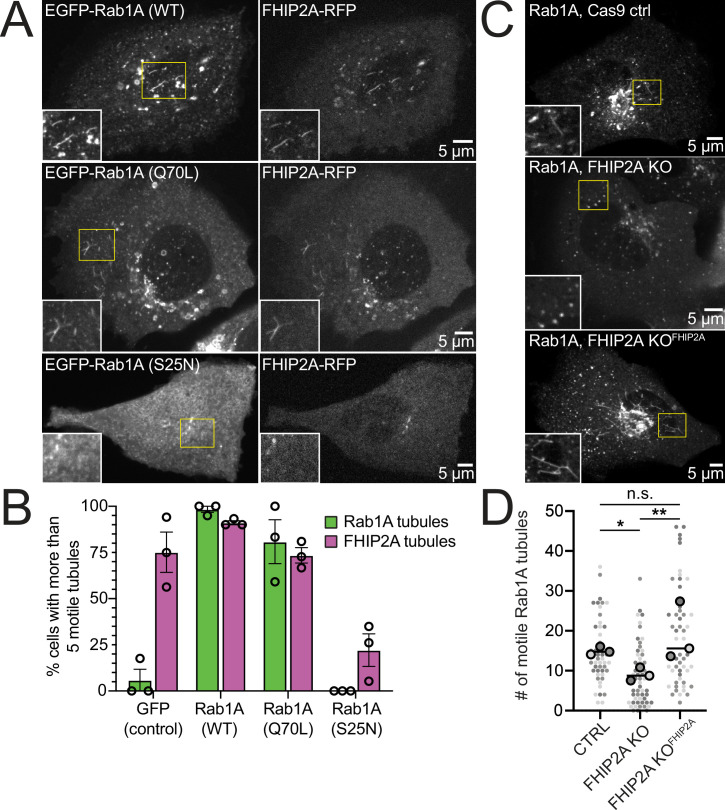
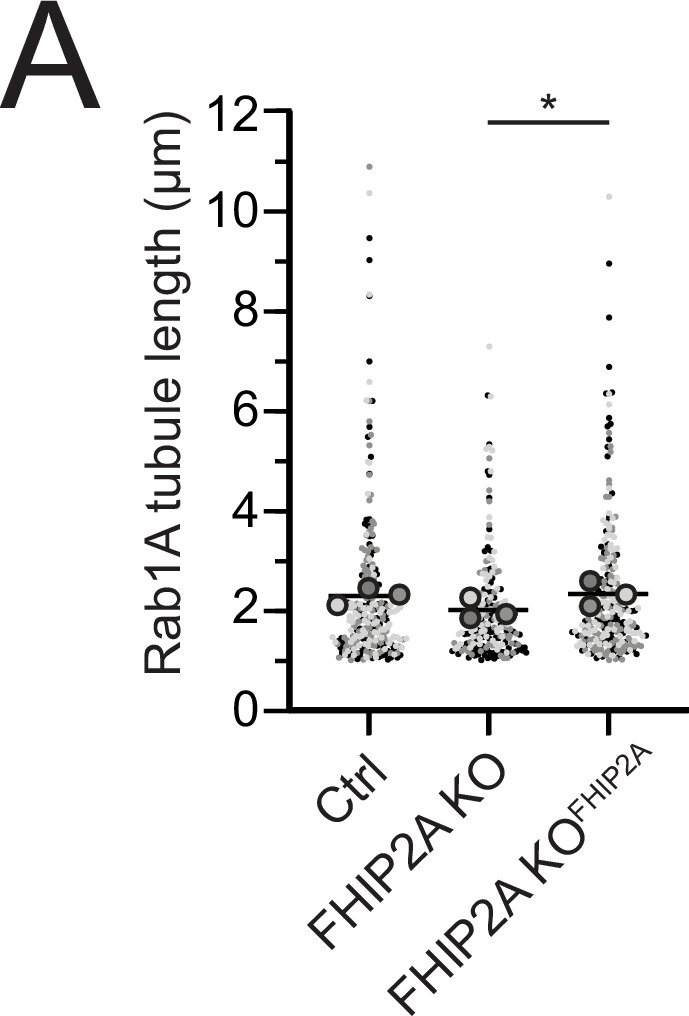
In eukaryotic cells, intracellular components are organized by the microtubule motors cytoplasmic dynein-1 (dynein) and kinesins, which are linked to cargos via adaptor proteins. While ~40 kinesins transport cargo toward the plus end of microtubules, a single dynein moves cargo in the opposite direction. How dynein transports a wide variety of cargos remains an open question. The FTS-Hook-FHIP ('FHF') cargo adaptor complex links dynein to cargo in humans and fungi. As human cells have three Hooks and four FHIP proteins, we hypothesized that the combinatorial assembly of different Hook and FHIP proteins could underlie dynein cargo diversity. Using proteomic approaches, we determine the protein 'interactome' of each FHIP protein. Live-cell imaging and biochemical approaches show that different FHF complexes associate with distinct motile cargos. These complexes also move with dynein and its cofactor dynactin in single-molecule in vitro reconstitution assays. Complexes composed of FTS, FHIP1B, and Hook1/Hook3 colocalize with Rab5-tagged early endosomes via a direct interaction between FHIP1B and GTP-bound Rab5. In contrast, complexes composed of FTS, FHIP2A, and Hook2 colocalize with Rab1A-tagged ER-to-Golgi cargos and FHIP2A is involved in the motility of Rab1A tubules. Our findings suggest that combinatorial assembly of different FTS-Hook-FHIP complexes is one mechanism dynein uses to achieve cargo specificity.
在真核细胞中,细胞内成分通过微管马达细胞质动力蛋白-1(dynein)和驱动蛋白来组织,这些马达通过衔接蛋白与货物相连。虽然大约有 40 种驱动蛋白将货物向微管的正极运输,但只有一种动力蛋白将货物向相反方向运输。动力蛋白如何运输各种各样的货物仍然是一个悬而未决的问题。FTS-Hook-FHIP(“FHF”)货物衔接复合物将 dynein 与人和真菌中的货物连接起来。由于人类细胞有三个 Hooks 和四个 FHIP 蛋白,我们假设不同 Hook 和 FHIP 蛋白的组合组装可能是 dynein 货物多样性的基础。我们使用蛋白质组学方法来确定每个 FHIP 蛋白的蛋白质“相互作用组”。活细胞成像和生化方法表明,不同的 FHF 复合物与不同的运动货物结合。这些复合物还在体外单分子重建测定中与 dynein 及其共因子 dynactin 一起移动。由 FTS、FHIP1B 和 Hook1/Hook3 组成的复合物通过 FHIP1B 和 GTP 结合的 Rab5 之间的直接相互作用与 Rab5 标记的早期内体共定位。相比之下,由 FTS、FHIP2A 和 Hook2 组成的复合物与 Rab1A 标记的 ER 到高尔基体货物共定位,并且 FHIP2A 参与 Rab1A 小管的运动。我们的研究结果表明,不同的 FTS-Hook-FHIP 复合物的组合组装是 dynein 实现货物特异性的一种机制。