CRUK Beatson Institute, Garscube Estate, Glasgow, United Kingdom.
Institute of Cancer Sciences, University of Glasgow, Glasgow, United Kingdom.
Elife. 2021 Dec 13;10:e69729. doi: 10.7554/eLife.69729.
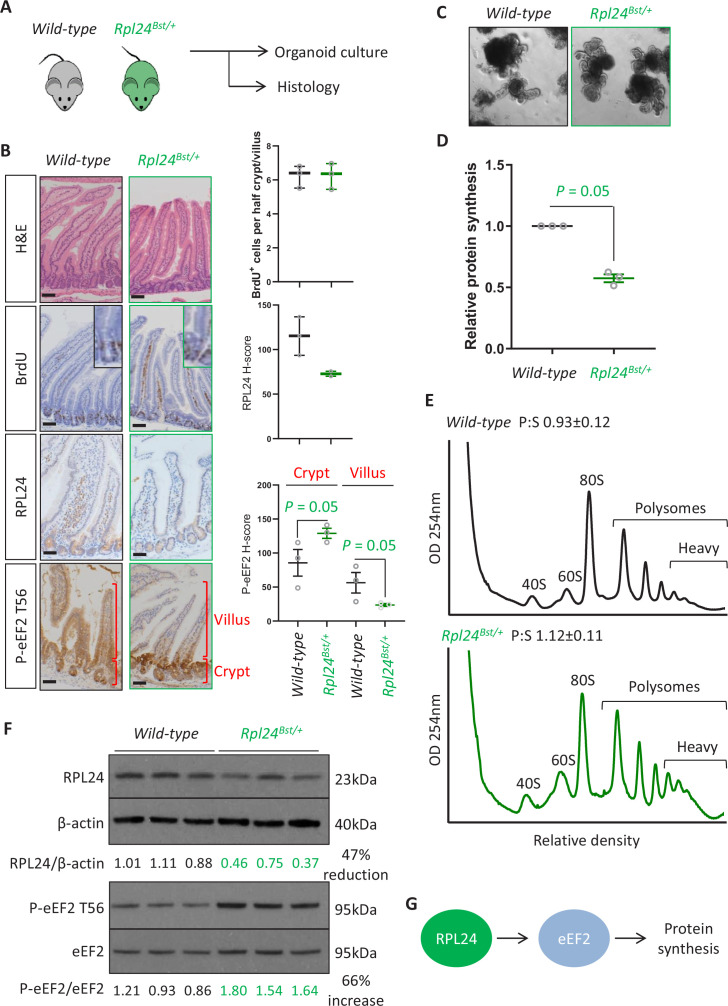
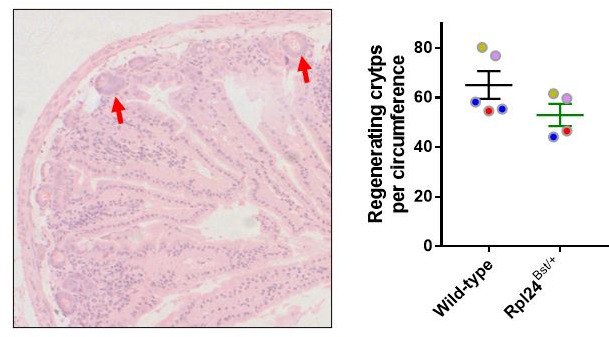
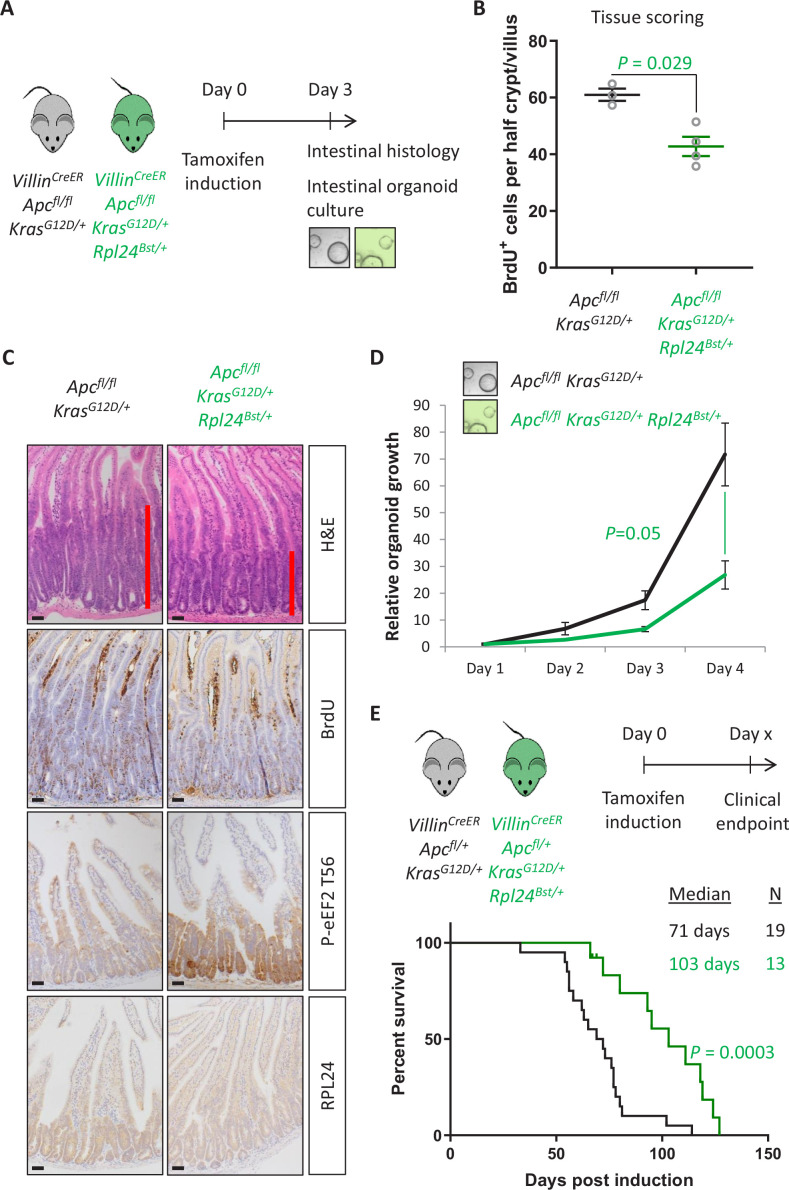
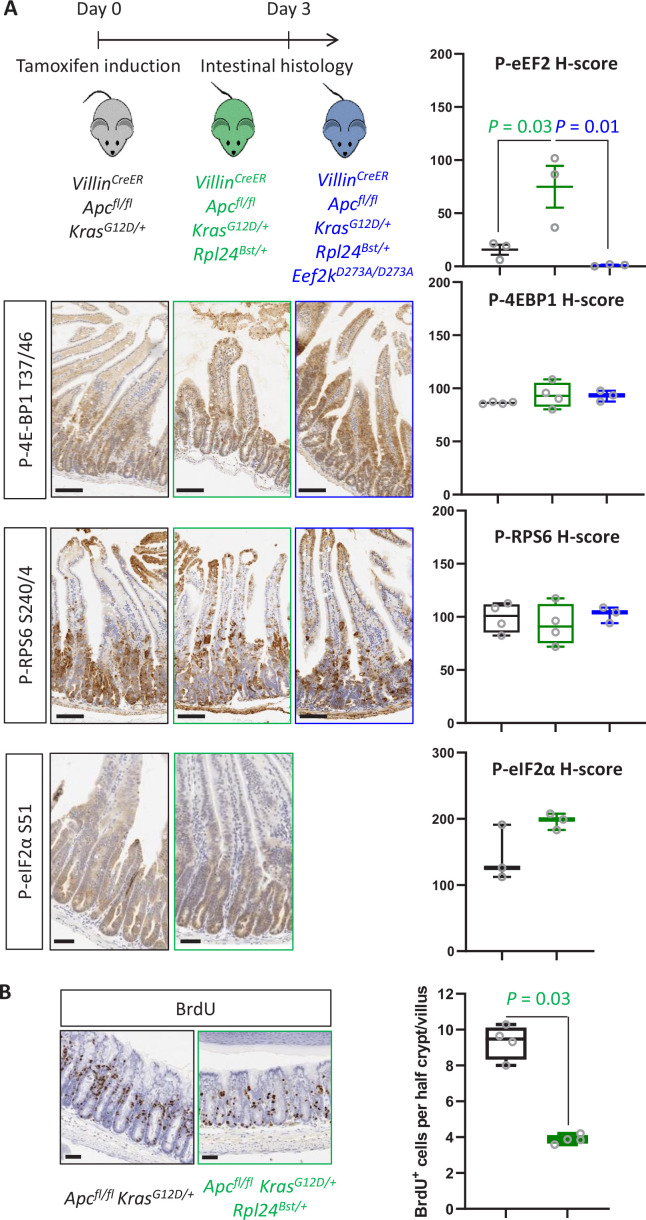
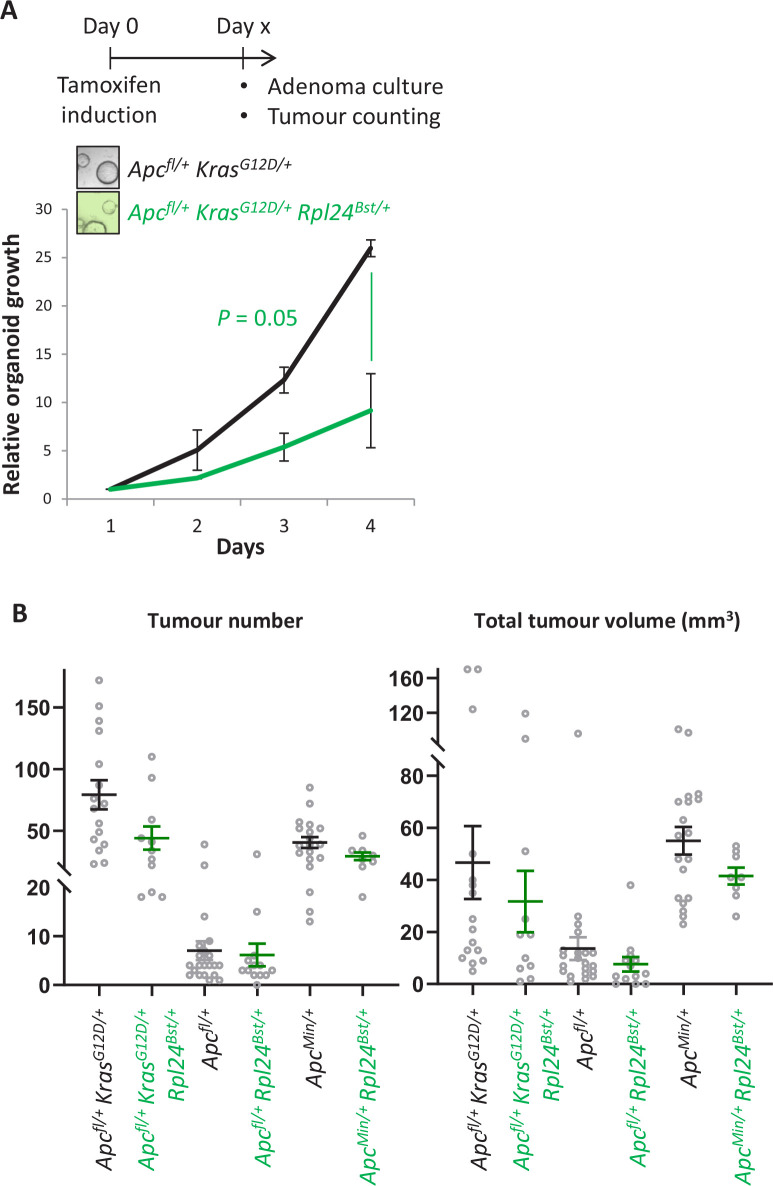
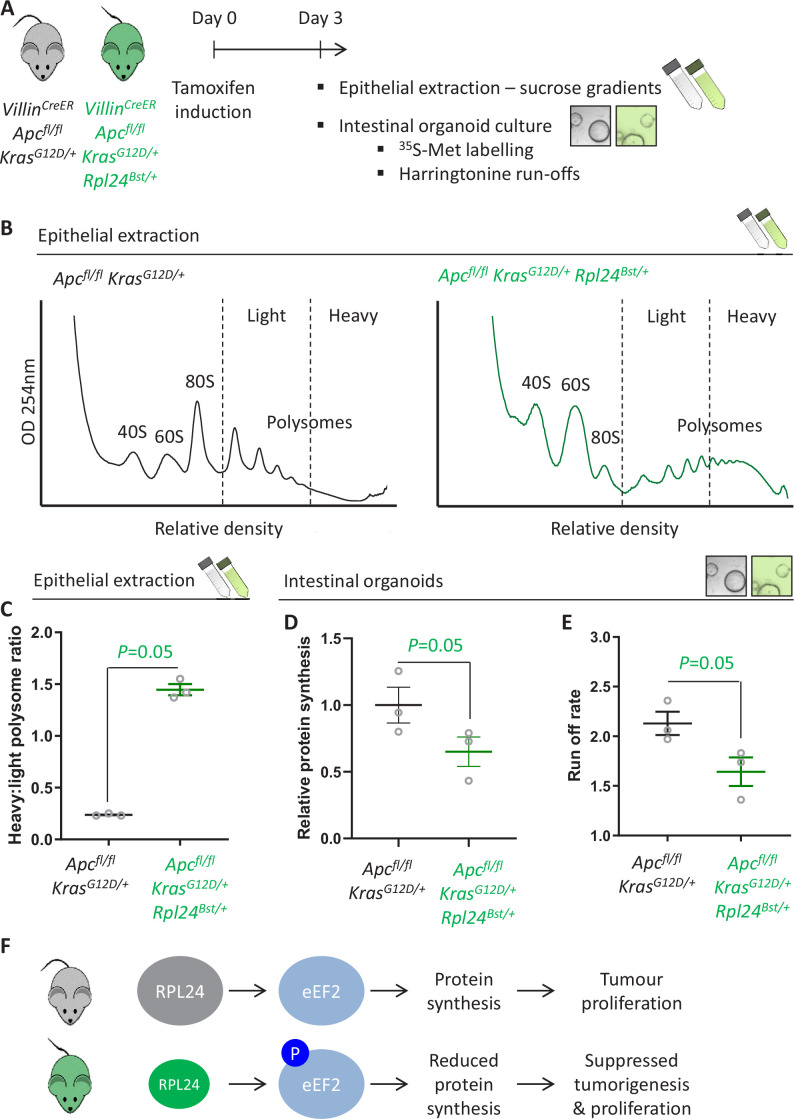
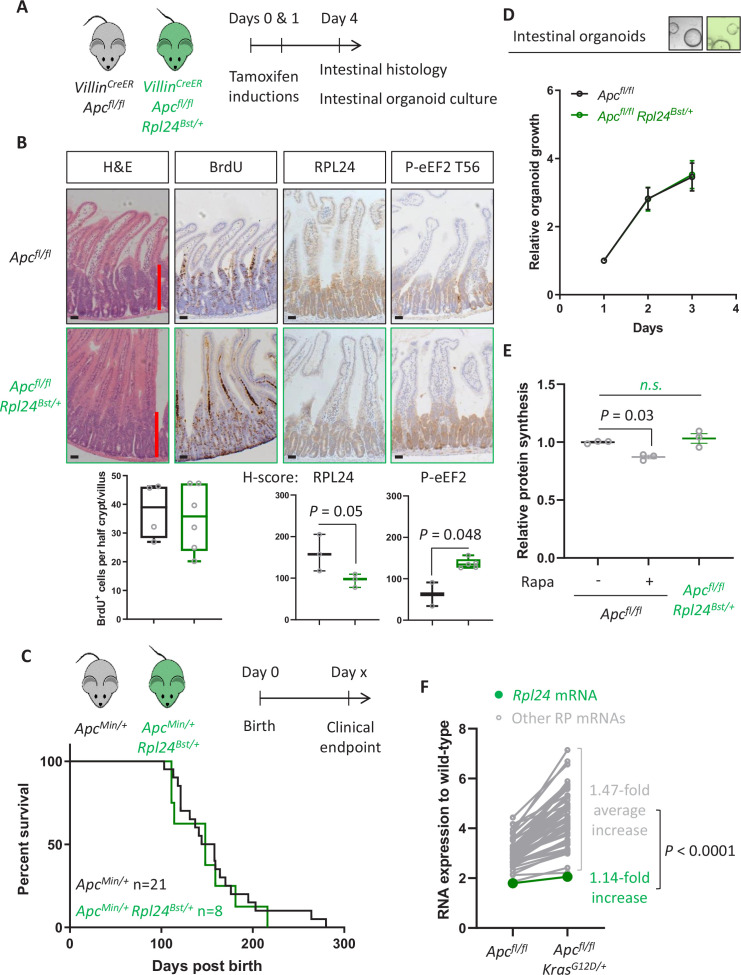
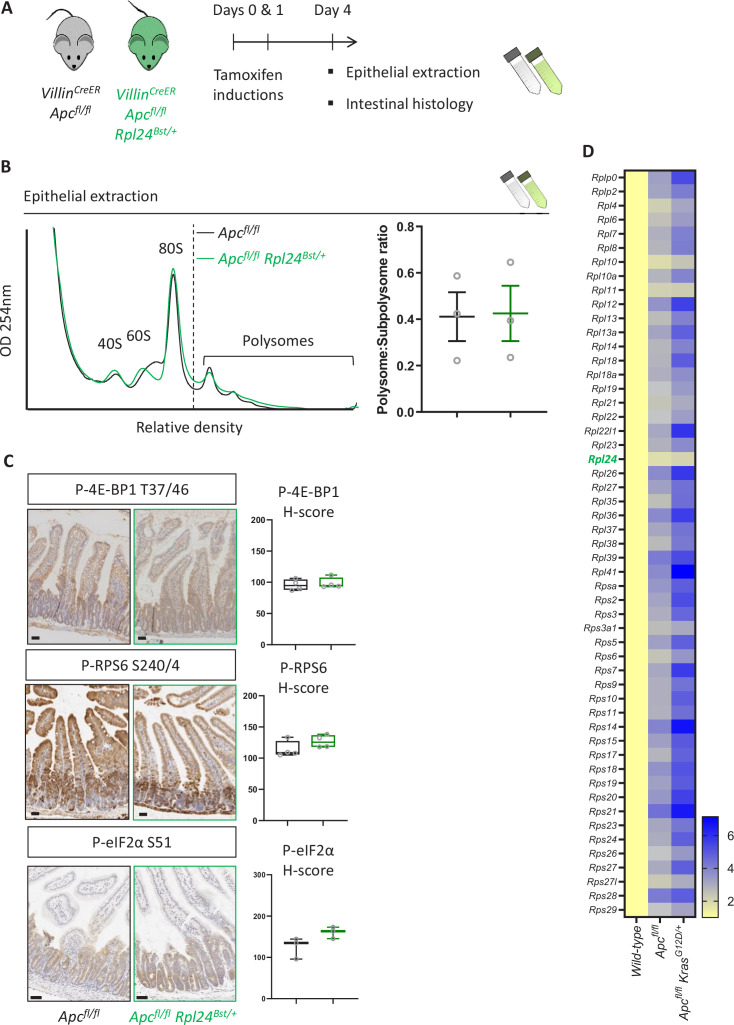
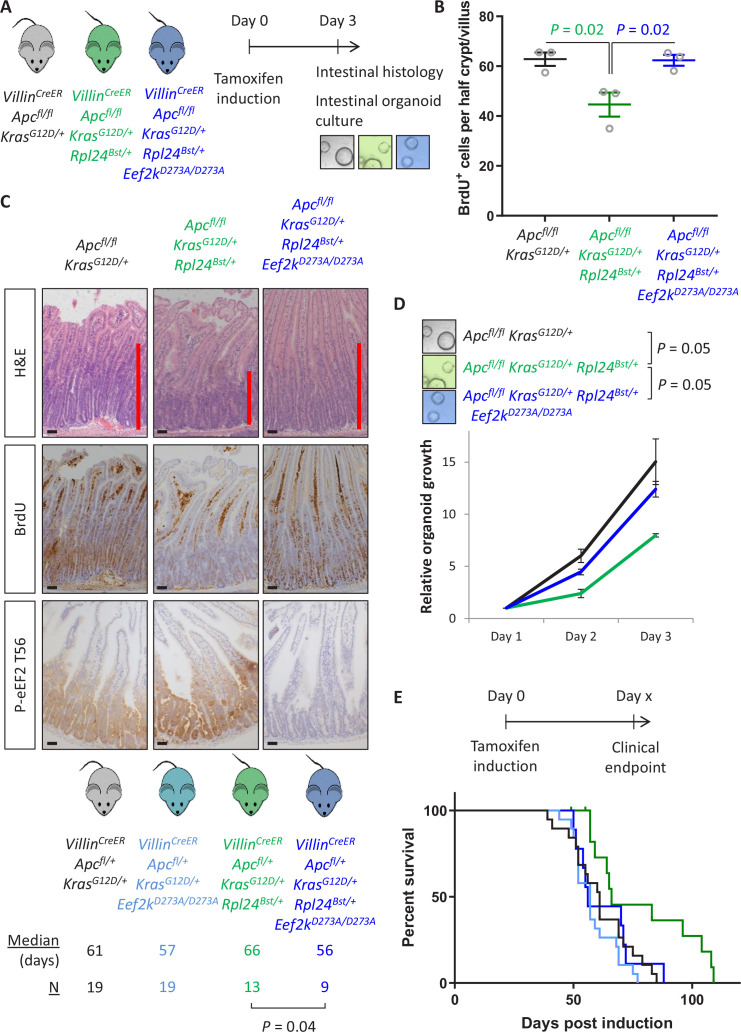
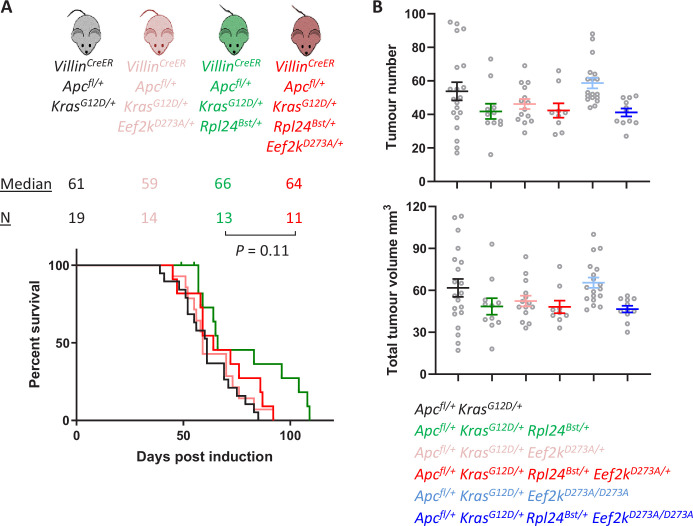
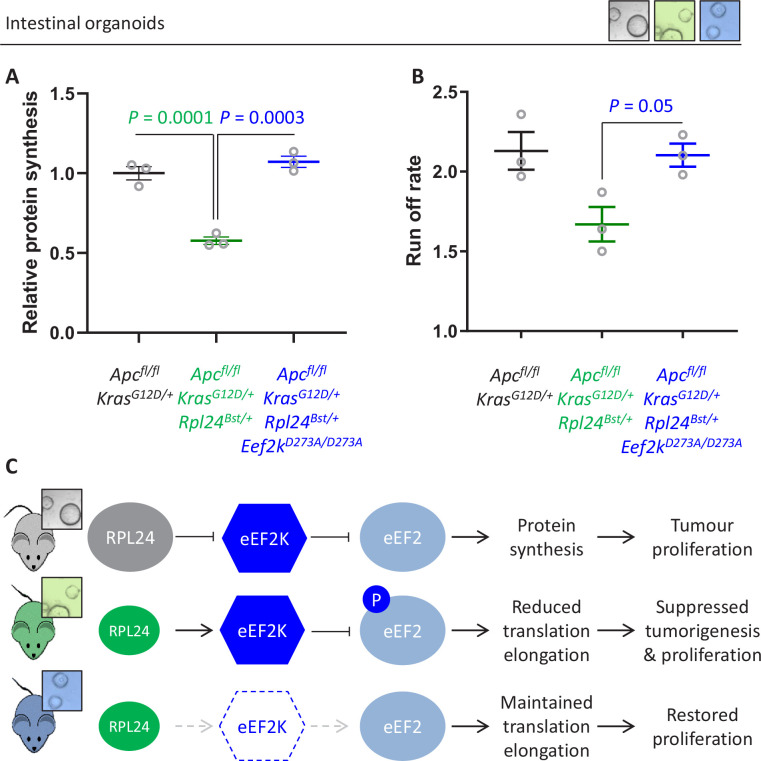
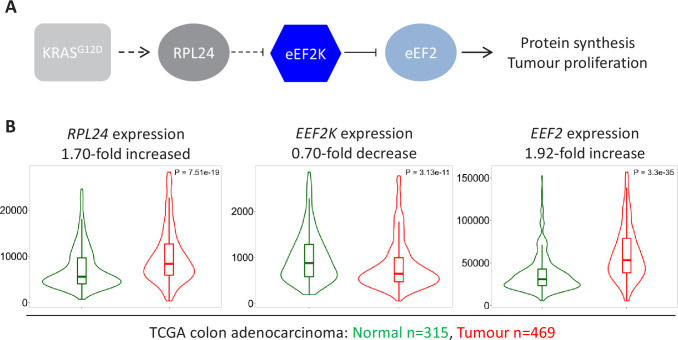
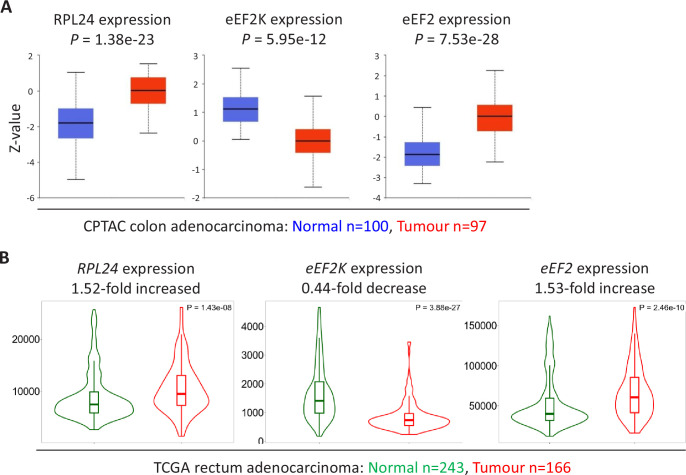
Increased protein synthesis supports the rapid cell proliferation associated with cancer. The mutant mouse reduces the expression of the ribosomal protein RPL24 and has been used to suppress translation and limit tumorigenesis in multiple mouse models of cancer. Here, we show that also suppresses tumorigenesis and proliferation in a model of colorectal cancer (CRC) with two common patient mutations, and . In contrast to previous reports, mutation has no effect on ribosomal subunit abundance but suppresses translation elongation through phosphorylation of eEF2, reducing protein synthesis by 40% in tumour cells. Ablating eEF2 phosphorylation in mutant mice by inactivating its kinase, eEF2K, completely restores the rates of elongation and protein synthesis. Furthermore, eEF2K activity is required for the mutant to suppress tumorigenesis. This work demonstrates that elevation of eEF2 phosphorylation is an effective means to suppress colorectal tumorigenesis with two driver mutations. This positions translation elongation as a therapeutic target in CRC, as well as in other cancers where the mutation has a tumour suppressive effect in mouse models.
蛋白质合成的增加支持与癌症相关的快速细胞增殖。RPL24 核糖体蛋白的突变小鼠减少了表达,并已被用于抑制翻译并限制多种癌症小鼠模型中的肿瘤发生。在这里,我们表明,也抑制了具有两种常见患者突变的结直肠癌(CRC)模型中的肿瘤发生和增殖,和。与之前的报告相反,突变对核糖体亚基丰度没有影响,但通过磷酸化 eEF2 抑制翻译延伸,使肿瘤细胞中的蛋白质合成减少 40%。通过使激酶 eEF2K 失活来消除 突变小鼠中的 eEF2 磷酸化,完全恢复延伸和蛋白质合成的速率。此外,eEF2K 活性对于 突变体抑制肿瘤发生是必需的。这项工作表明,升高的 eEF2 磷酸化是抑制具有两种驱动突变的结直肠肿瘤发生的有效方法。这将翻译延伸定位为 CRC 以及其他在小鼠模型中具有肿瘤抑制作用的癌症的治疗靶点。