Aalto Acoustics Lab, Department of Signal Processing and Acoustics, School of Electrical Engineering, Aalto University, Espoo, Finland.
J Assoc Res Otolaryngol. 2022 Feb;23(1):75-94. doi: 10.1007/s10162-021-00820-4. Epub 2021 Dec 13.
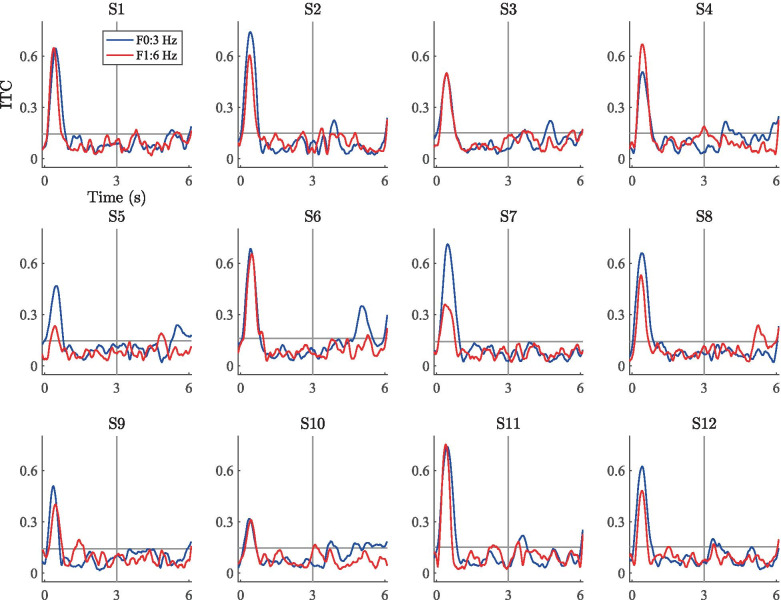
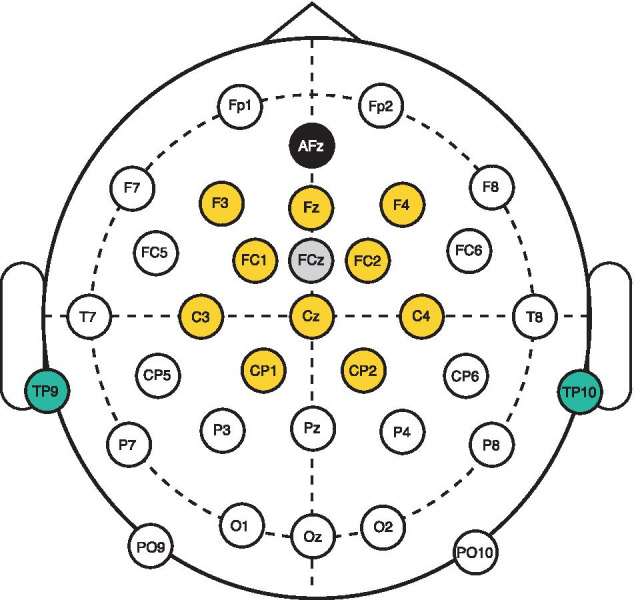
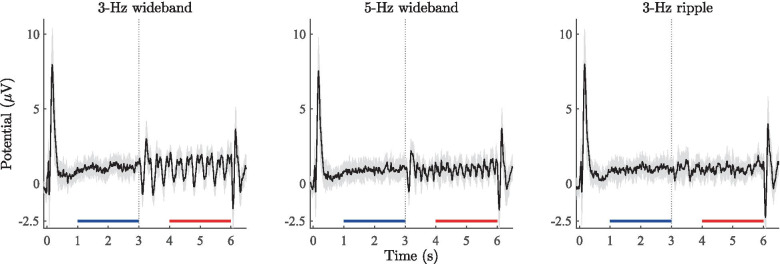
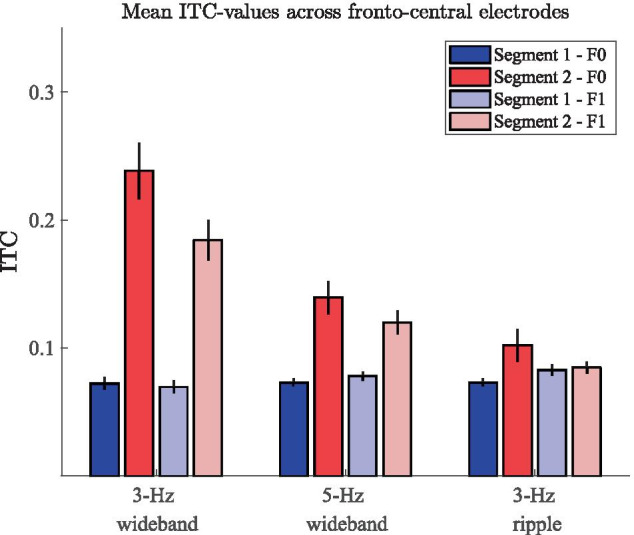
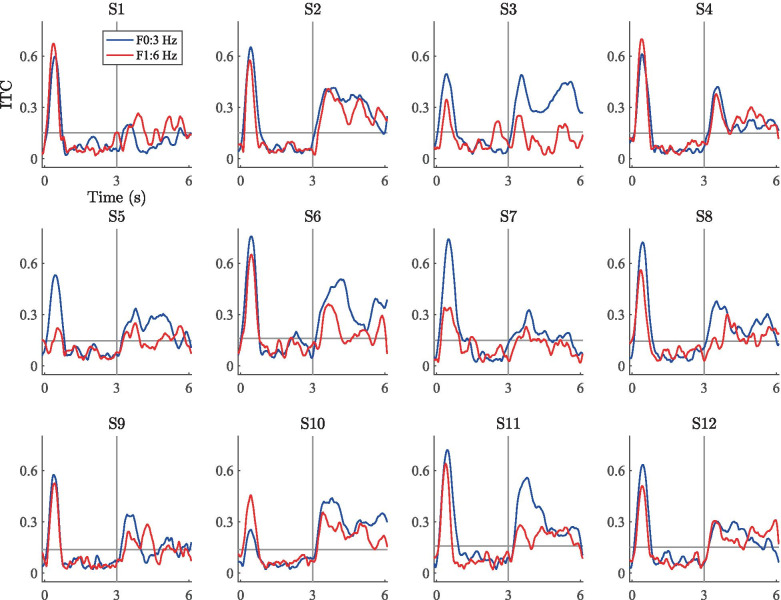
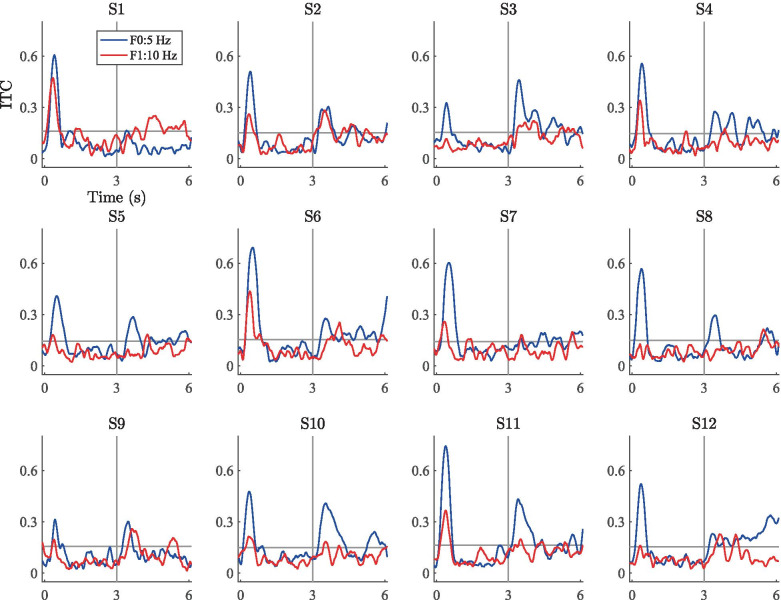
Spatial hearing facilitates the perceptual organization of complex soundscapes into accurate mental representations of sound sources in the environment. Yet, the role of binaural cues in auditory scene analysis (ASA) has received relatively little attention in recent neuroscientific studies employing novel, spectro-temporally complex stimuli. This may be because a stimulation paradigm that provides binaurally derived grouping cues of sufficient spectro-temporal complexity has not yet been established for neuroscientific ASA experiments. Random-chord stereograms (RCS) are a class of auditory stimuli that exploit spectro-temporal variations in the interaural envelope correlation of noise-like sounds with interaurally coherent fine structure; they evoke salient auditory percepts that emerge only under binaural listening. Here, our aim was to assess the usability of the RCS paradigm for indexing binaural processing in the human brain. To this end, we recorded EEG responses to RCS stimuli from 12 normal-hearing subjects. The stimuli consisted of an initial 3-s noise segment with interaurally uncorrelated envelopes, followed by another 3-s segment, where envelope correlation was modulated periodically according to the RCS paradigm. Modulations were applied either across the entire stimulus bandwidth (wideband stimuli) or in temporally shifting frequency bands (ripple stimulus). Event-related potentials and inter-trial phase coherence analyses of the EEG responses showed that the introduction of the 3- or 5-Hz wideband modulations produced a prominent change-onset complex and ongoing synchronized responses to the RCS modulations. In contrast, the ripple stimulus elicited a change-onset response but no response to ongoing RCS modulation. Frequency-domain analyses revealed increased spectral power at the fundamental frequency and the first harmonic of wideband RCS modulations. RCS stimulation yields robust EEG measures of binaurally driven auditory reorganization and has potential to provide a flexible stimulation paradigm suitable for isolating binaural effects in ASA experiments.
空间听觉有助于将复杂的声音环境感知组织为环境中声源的准确心理表示。然而,在最近采用新颖的、具有复杂时频谱的刺激进行的神经科学研究中,双耳线索在听觉场景分析 (ASA) 中的作用相对较少受到关注。这可能是因为尚未为神经科学 ASA 实验建立提供足够复杂的时频谱双耳分组线索的刺激范式。随机弦立体图 (RCS) 是一类听觉刺激,它们利用类似噪声声音的耳间包络相关中的时频谱变化,以及耳间相干精细结构;它们引起仅在双耳聆听下才出现的明显听觉感知。在这里,我们的目的是评估 RCS 范式在人类大脑中索引双耳处理的可用性。为此,我们从 12 名正常听力受试者记录了对 RCS 刺激的 EEG 反应。刺激由初始 3 秒噪声段组成,耳间包络不相关,然后是另一个 3 秒段,其中包络根据 RCS 范式周期性地调制。调制要么在整个刺激带宽(宽带刺激)中进行,要么在时间上转移的频带(波纹刺激)中进行。EEG 响应的事件相关电位和试验间相位相干性分析表明,引入 3 或 5 Hz 宽带调制会产生明显的起始复合和对 RCS 调制的持续同步响应。相比之下,波纹刺激会引起起始响应,但对持续的 RCS 调制没有响应。频域分析显示宽带 RCS 调制的基频和第一谐波的谱功率增加。RCS 刺激产生了双耳驱动听觉重组的强大 EEG 测量值,并且具有提供适合于在 ASA 实验中分离双耳效应的灵活刺激范式的潜力。