Department of Cellular and Molecular Pharmacology, University of California San Francisco, San Francisco, United States.
Department of Biomolecular Sciences, Weizmann Institute of Science, Rehovot, Israel.
Elife. 2022 Jan 11;11:e70017. doi: 10.7554/eLife.70017.
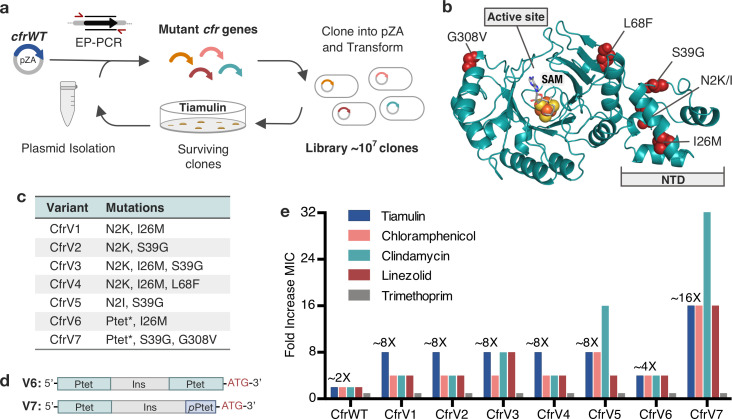
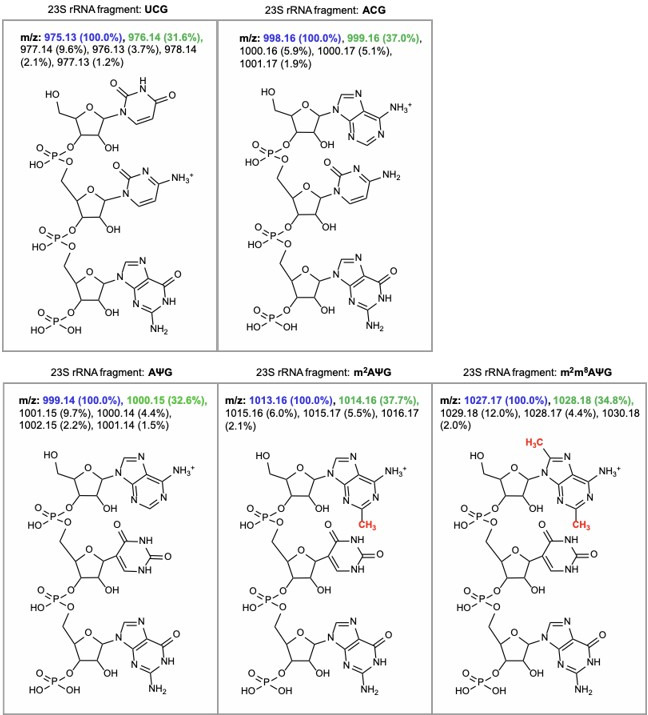
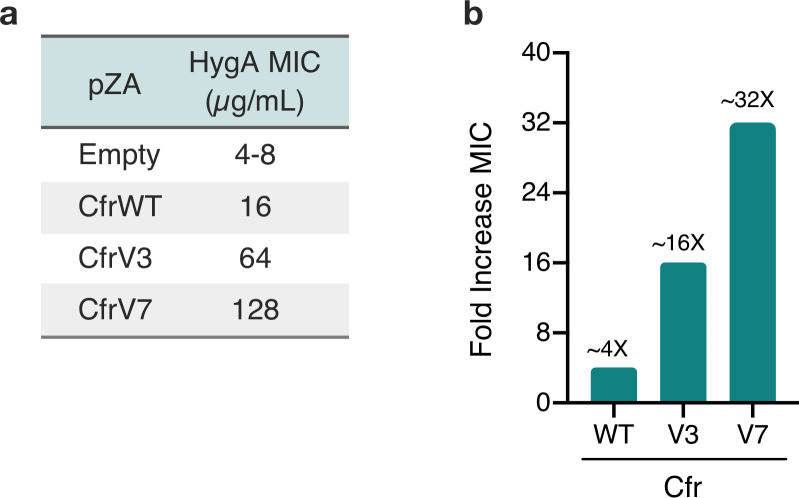
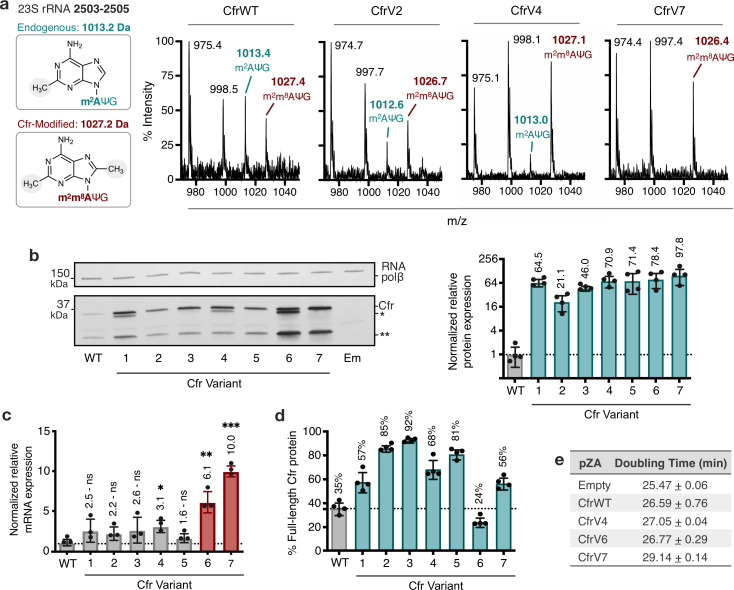
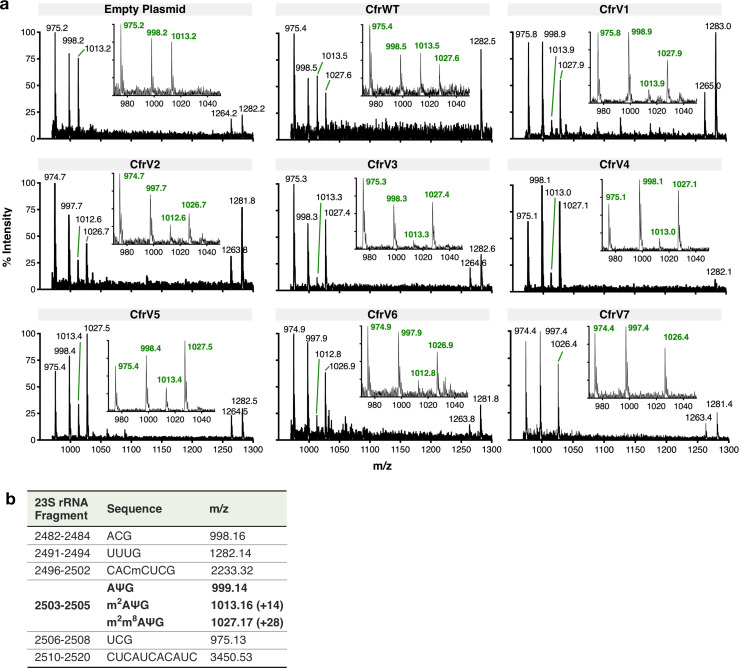
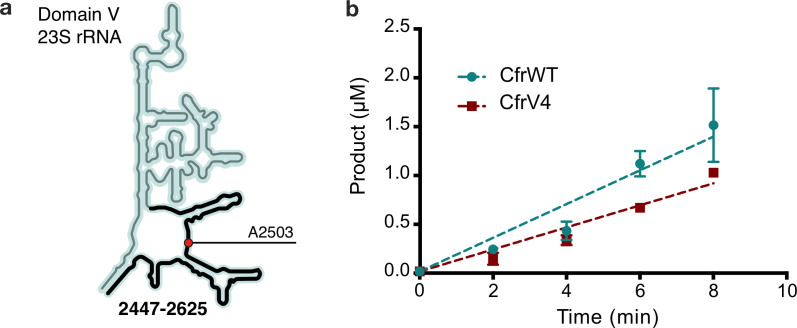
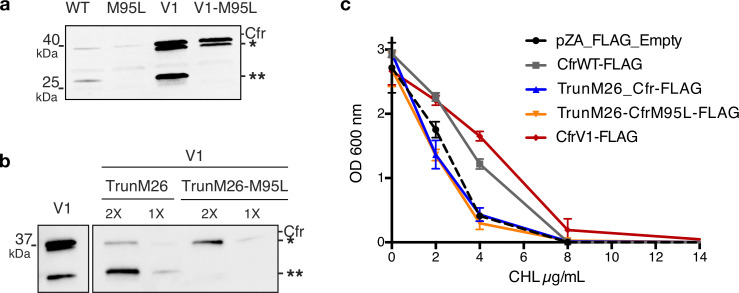
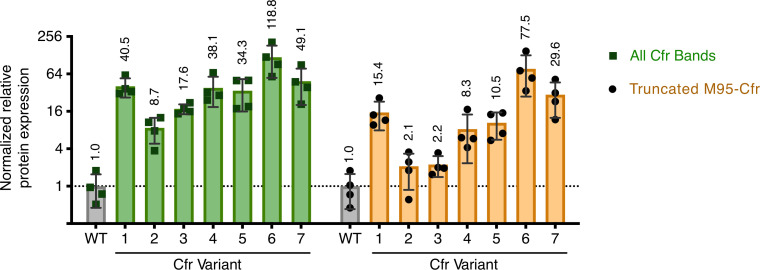
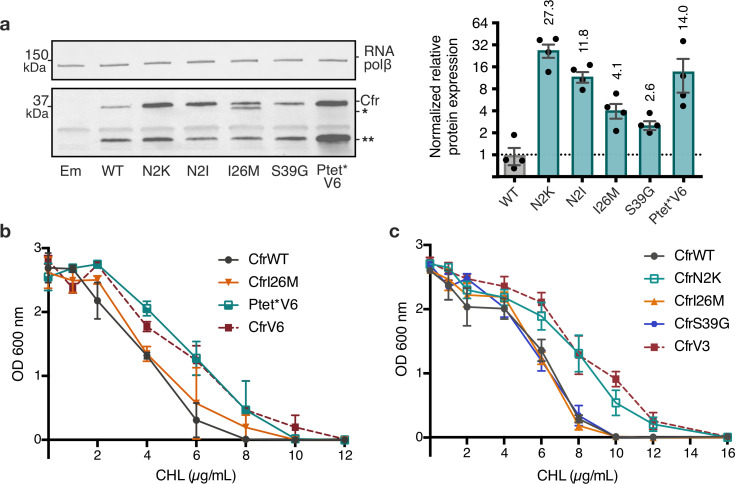
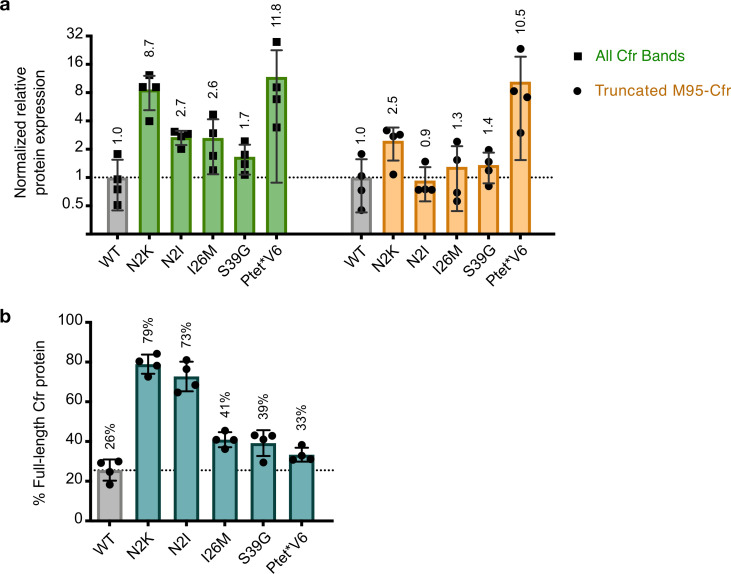
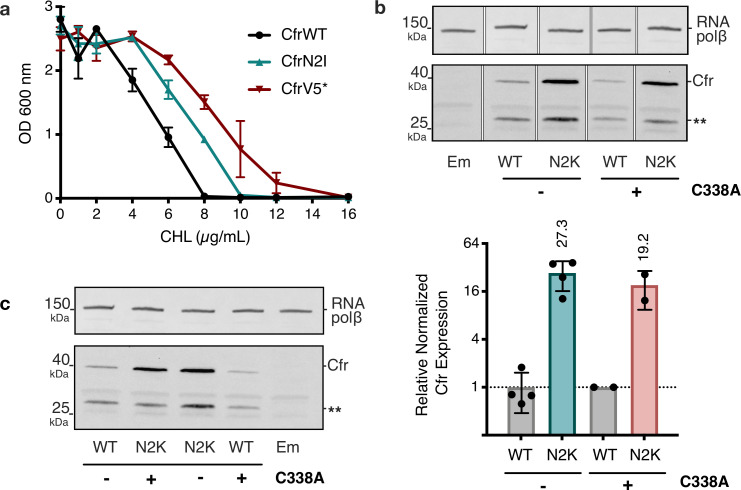
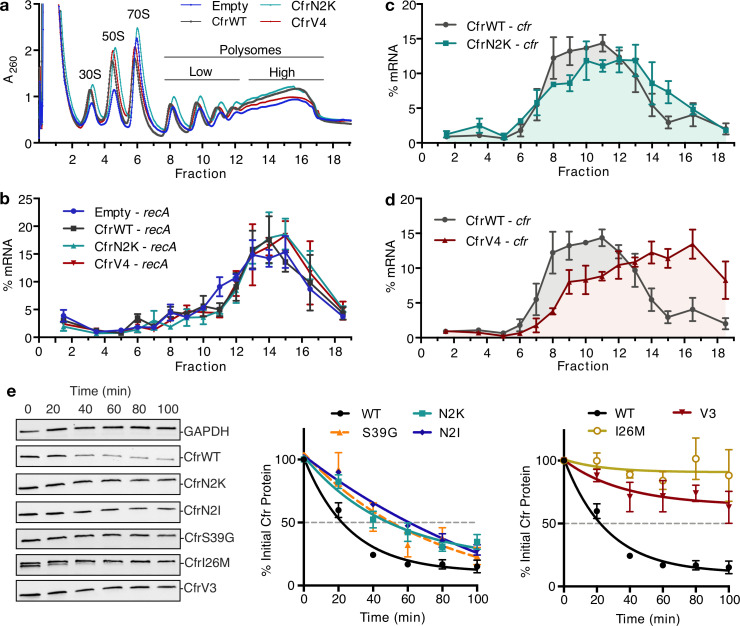
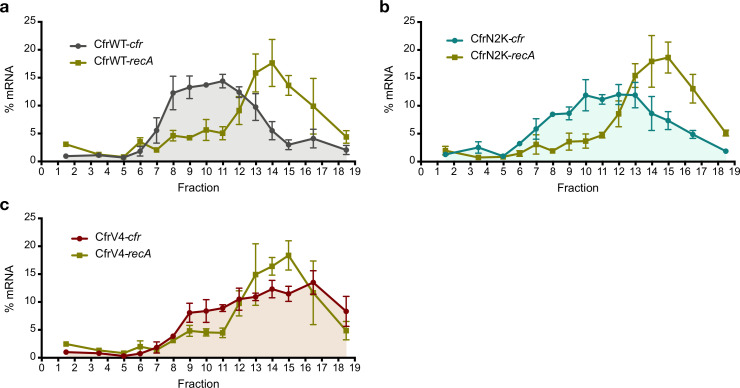
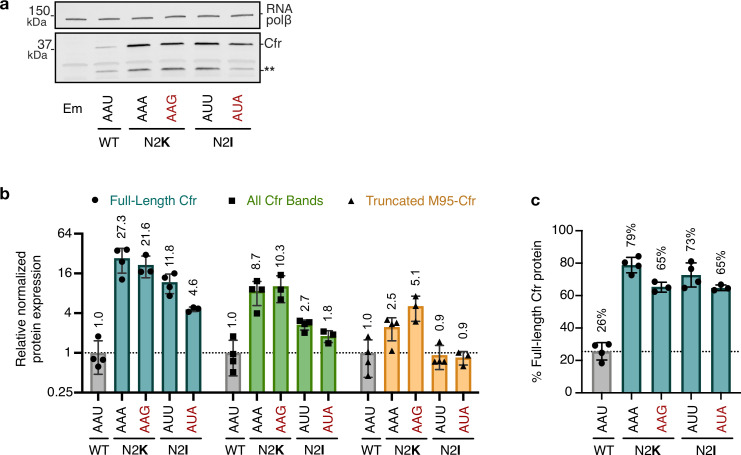
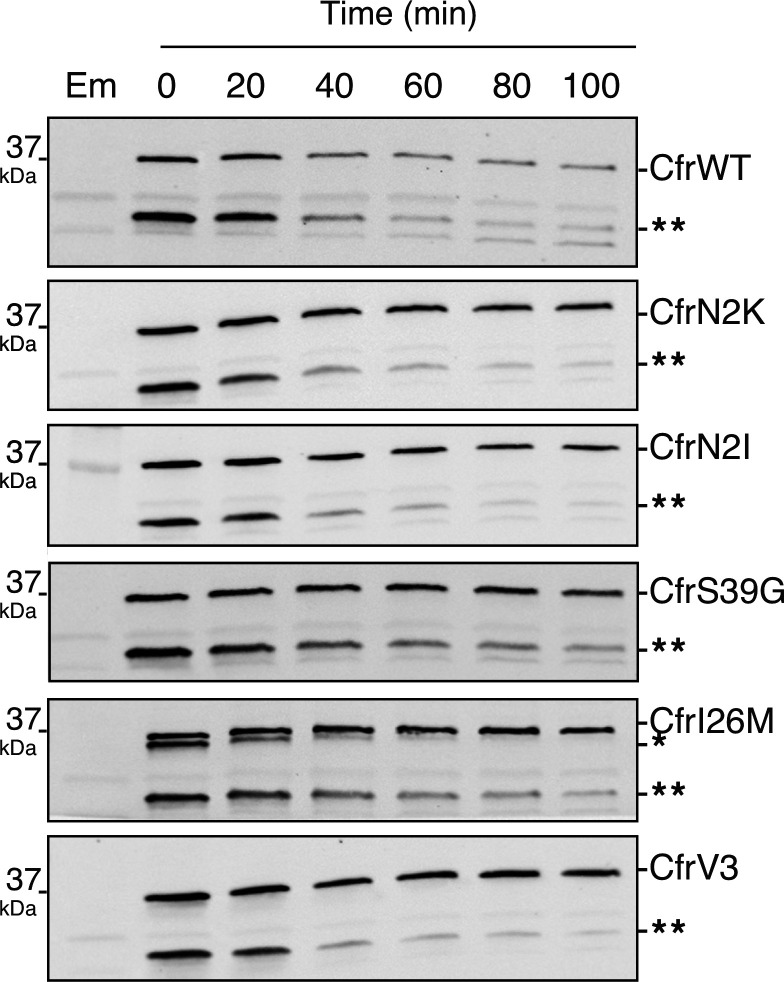
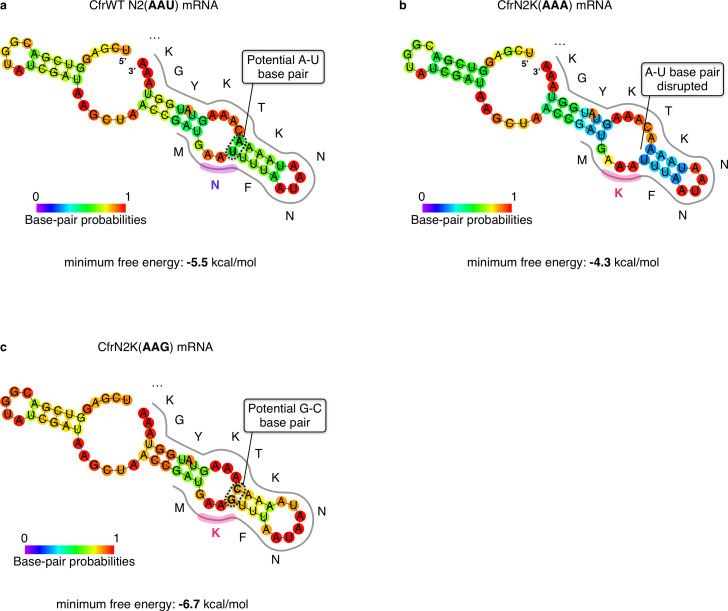
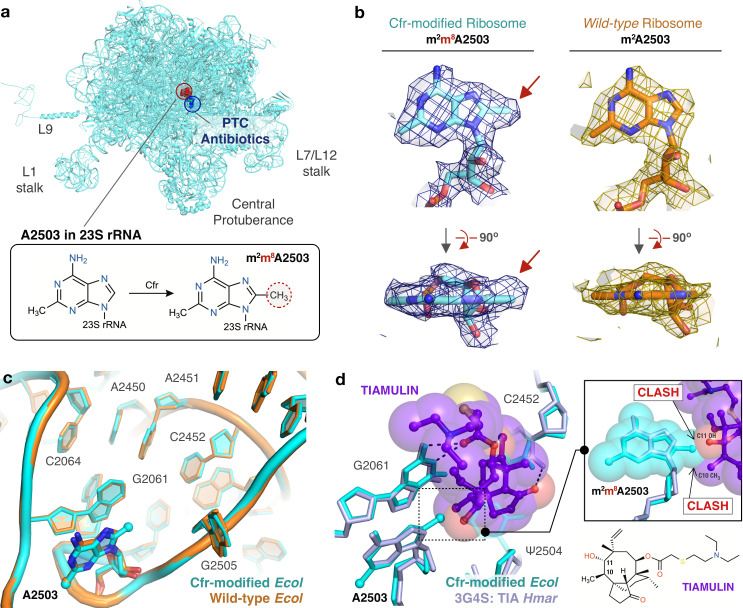
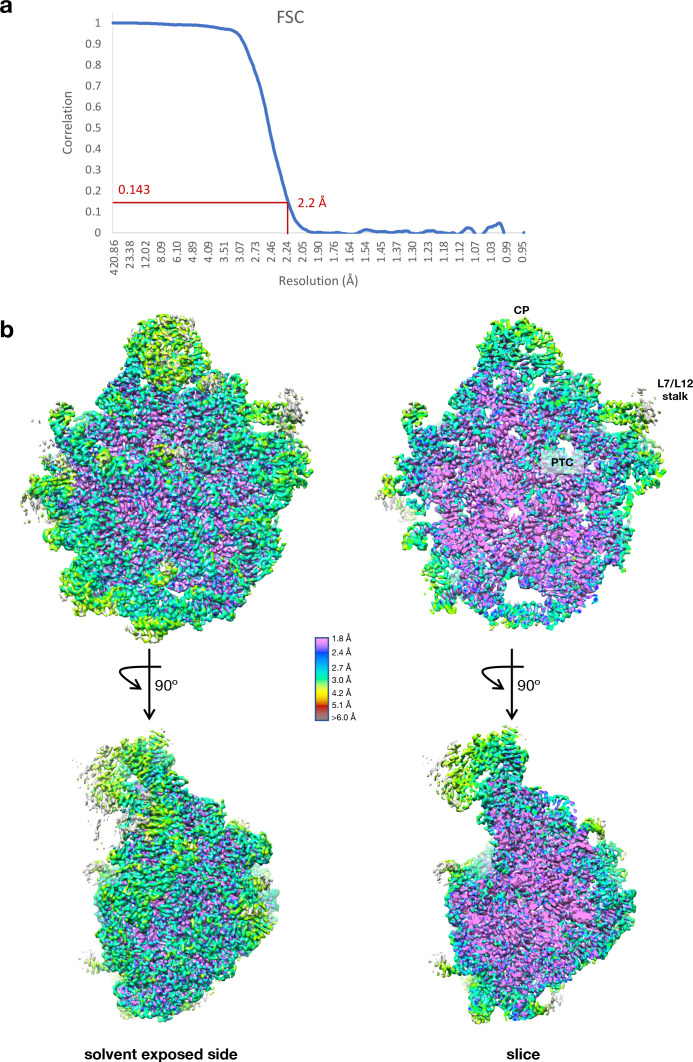
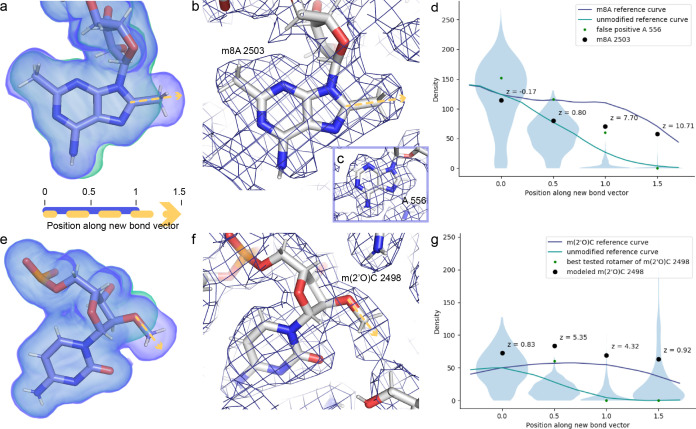
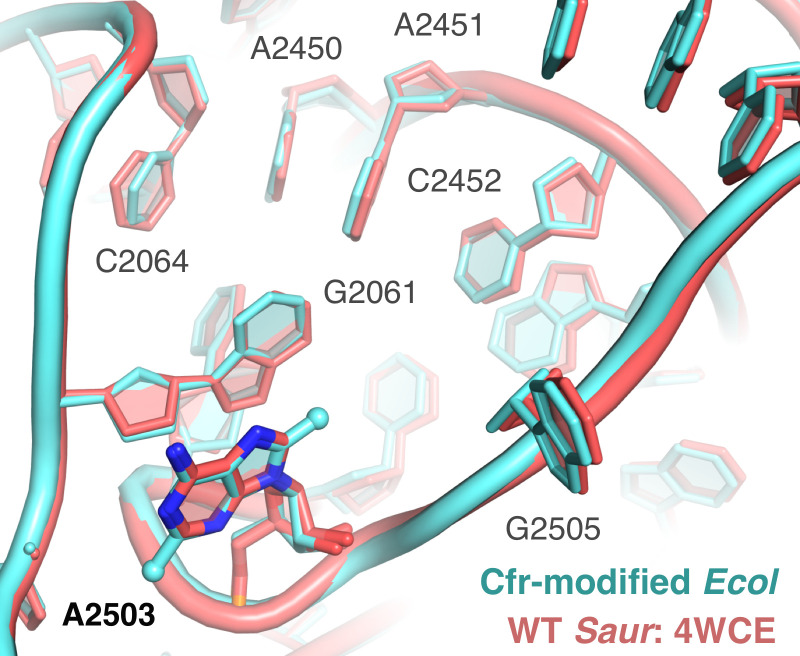
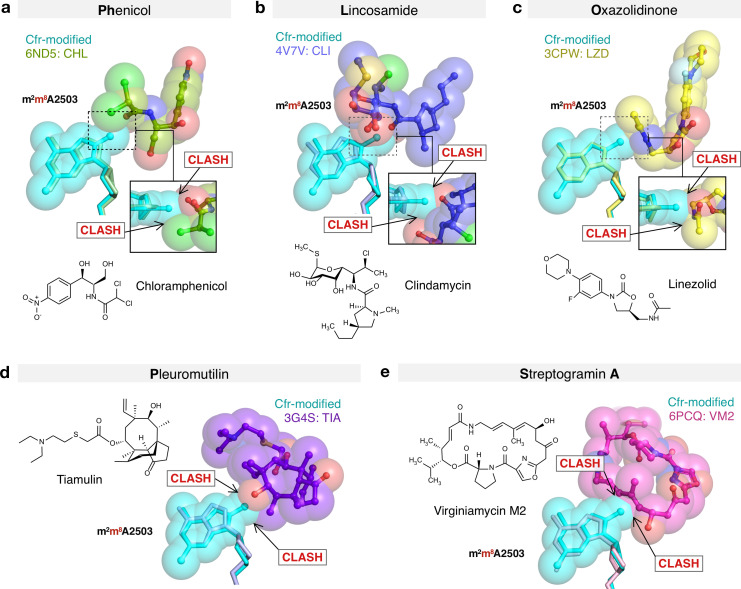
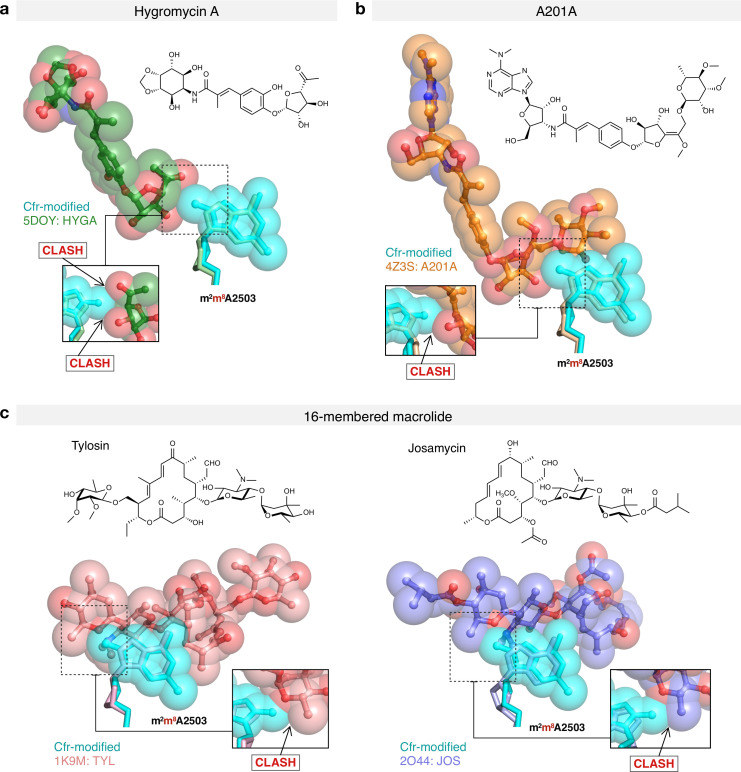
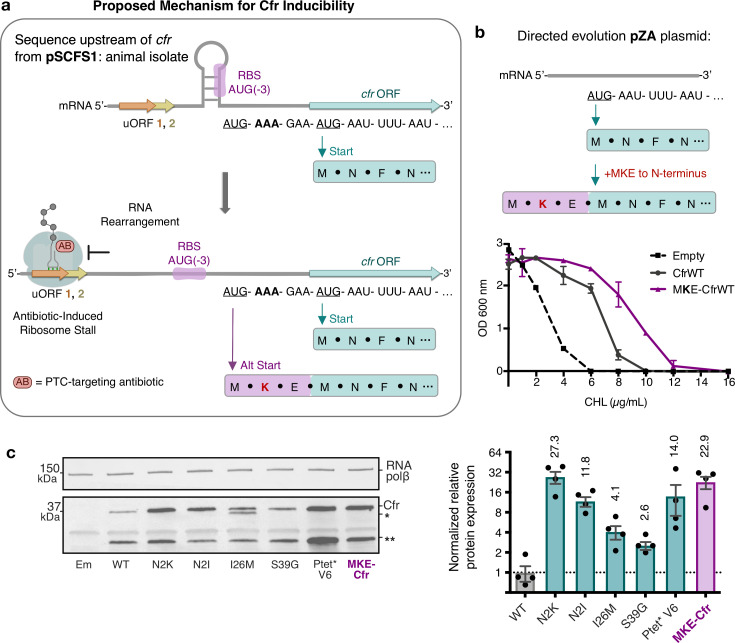
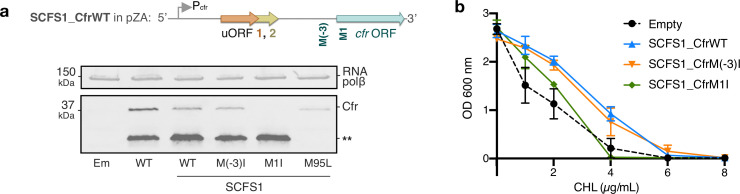
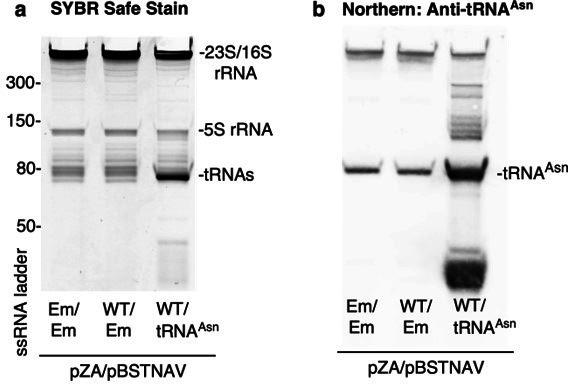
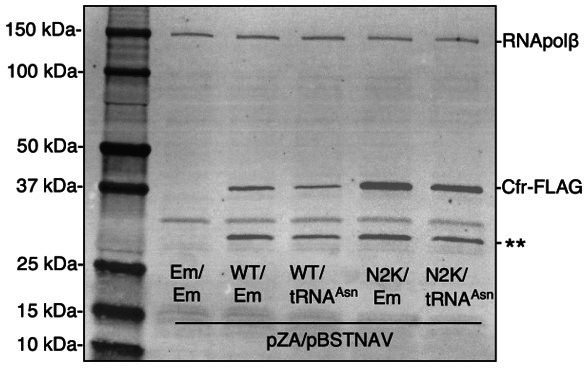
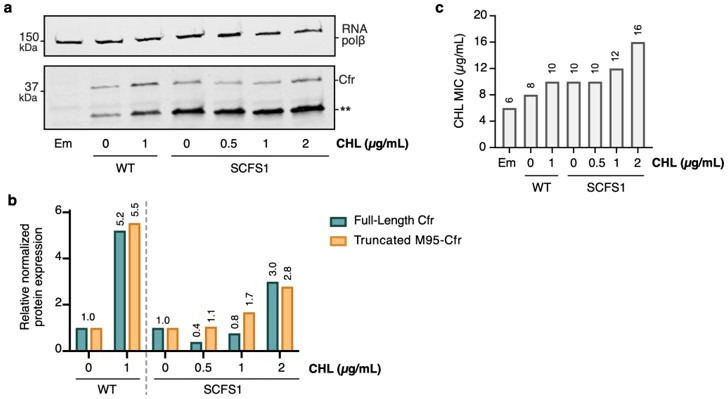
Alteration of antibiotic binding sites through modification of ribosomal RNA (rRNA) is a common form of resistance to ribosome-targeting antibiotics. The rRNA-modifying enzyme Cfr methylates an adenosine nucleotide within the peptidyl transferase center, resulting in the C-8 methylation of A2503 (mA2503). Acquisition of results in resistance to eight classes of ribosome-targeting antibiotics. Despite the prevalence of this resistance mechanism, it is poorly understood whether and how bacteria modulate Cfr methylation to adapt to antibiotic pressure. Moreover, direct evidence for how mA2503 alters antibiotic binding sites within the ribosome is lacking. In this study, we performed directed evolution of Cfr under antibiotic selection to generate Cfr variants that confer increased resistance by enhancing methylation of A2503 in cells. Increased rRNA methylation is achieved by improved expression and stability of Cfr through transcriptional and post-transcriptional mechanisms, which may be exploited by pathogens under antibiotic stress as suggested by natural isolates. Using a variant that achieves near-stoichiometric methylation of rRNA, we determined a 2.2 Å cryo-electron microscopy structure of the Cfr-modified ribosome. Our structure reveals the molecular basis for broad resistance to antibiotics and will inform the design of new antibiotics that overcome resistance mediated by Cfr.
核糖体 RNA(rRNA)结合位点的改变通过修饰核糖体 RNA 是针对核糖体抗生素的常见耐药形式。rRNA 修饰酶 Cfr 使肽基转移酶中心内的一个腺苷核苷酸甲基化,导致 A2503 的 C-8 甲基化(mA2503)。获得导致对八类核糖体靶向抗生素的耐药性。尽管这种耐药机制很常见,但人们对细菌是否以及如何调节 Cfr 甲基化以适应抗生素压力知之甚少。此外,缺乏 mA2503 如何改变核糖体中抗生素结合位点的直接证据。在这项研究中,我们在抗生素选择下对 Cfr 进行了定向进化,以产生通过增强细胞中 A2503 的甲基化来赋予更高抗性的 Cfr 变体。通过转录和转录后机制提高 Cfr 的表达和稳定性来实现 rRNA 甲基化的增加,这可能是病原体在抗生素压力下利用的,正如天然分离株所表明的那样。使用实现 rRNA 近乎化学计量甲基化的变体,我们确定了 Cfr 修饰核糖体的 2.2 Å 冷冻电镜结构。我们的结构揭示了广泛对抗生素耐药的分子基础,并将为设计克服由 Cfr 介导的耐药性的新抗生素提供信息。