Miura Masahiro C, Nagata Shohei, Tamaki Satoshi, Tomita Masaru, Kanai Akio
Institute for Advanced Biosciences, Keio University, Tsuruoka, Japan.
Systems Biology Program, Graduate School of Media and Governance, Keio University, Fujisawa, Japan.
Front Microbiol. 2022 Feb 28;13:849080. doi: 10.3389/fmicb.2022.849080. eCollection 2022.
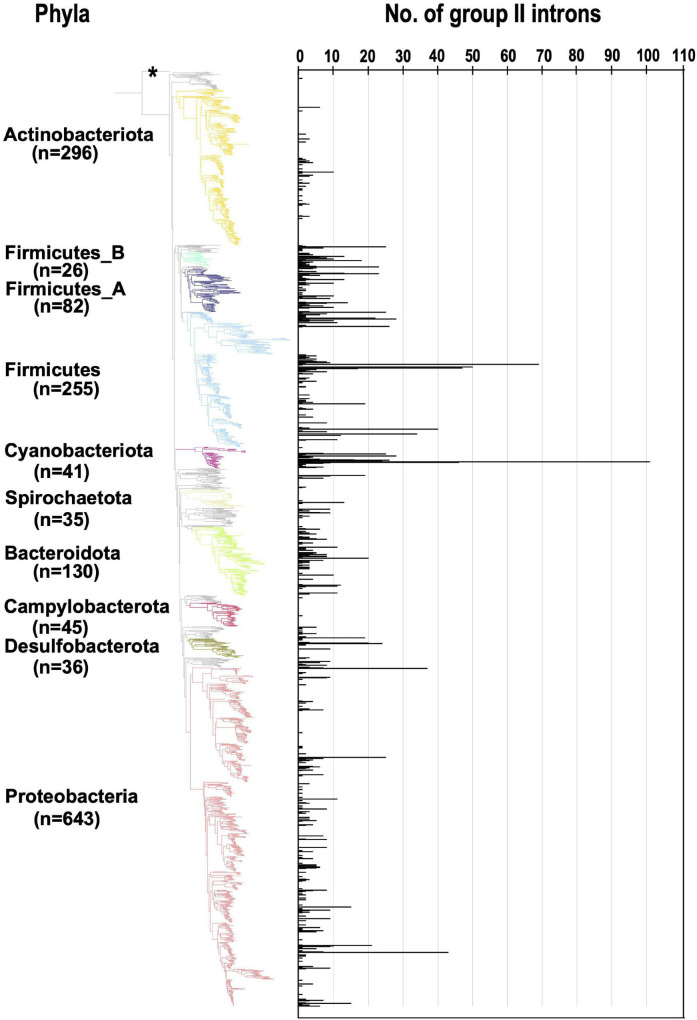
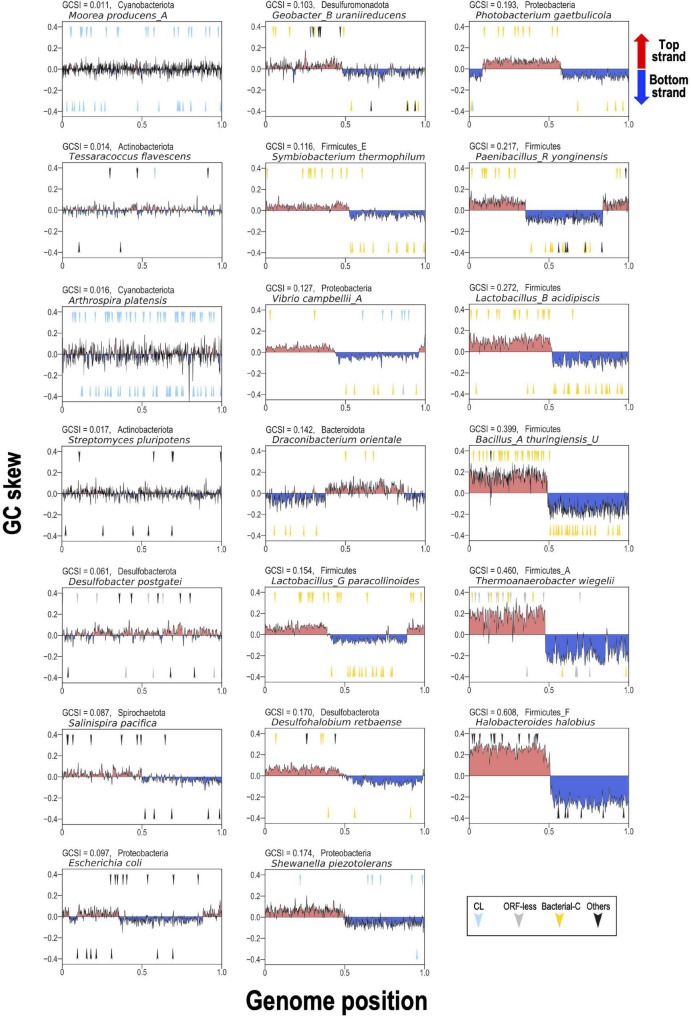
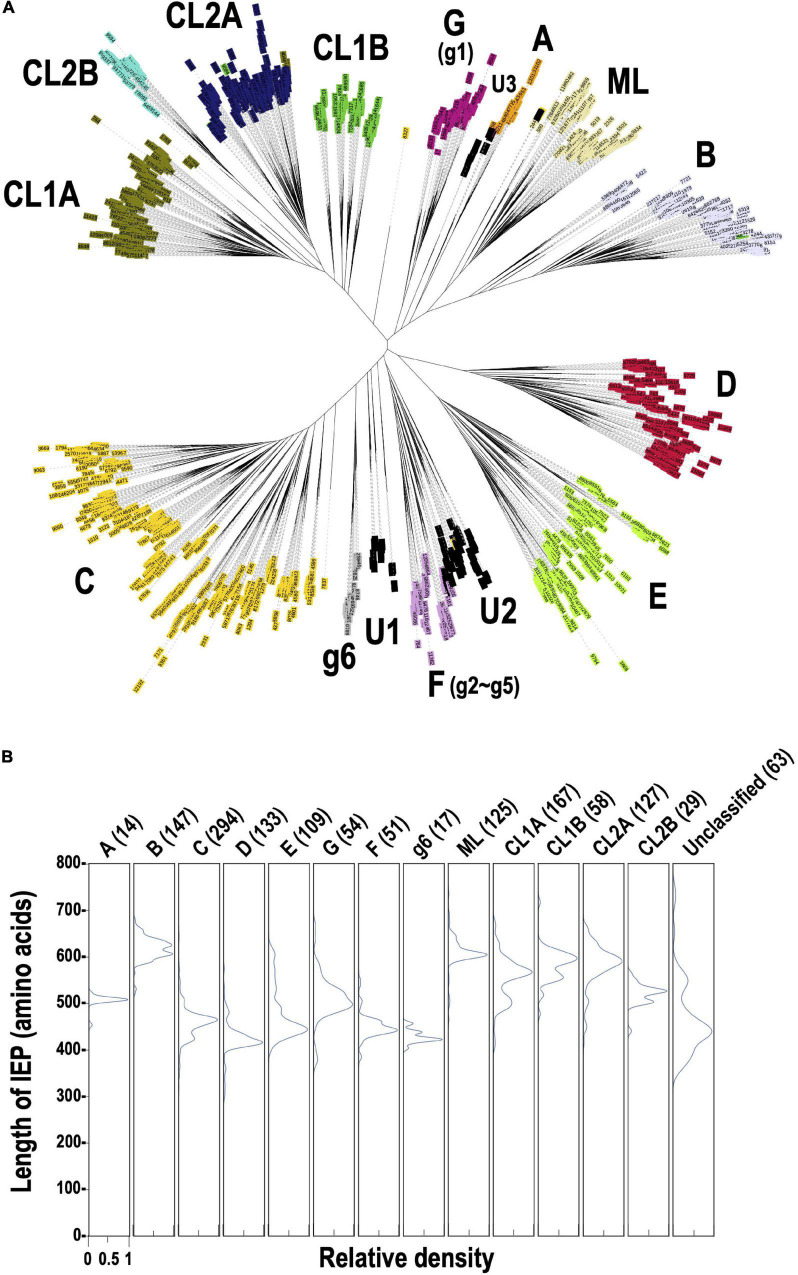
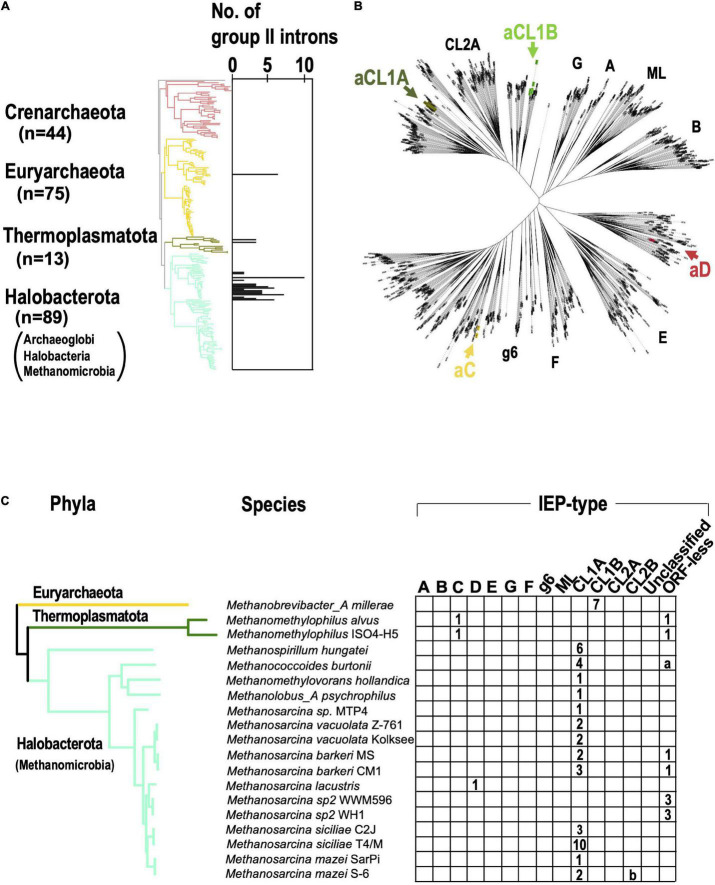
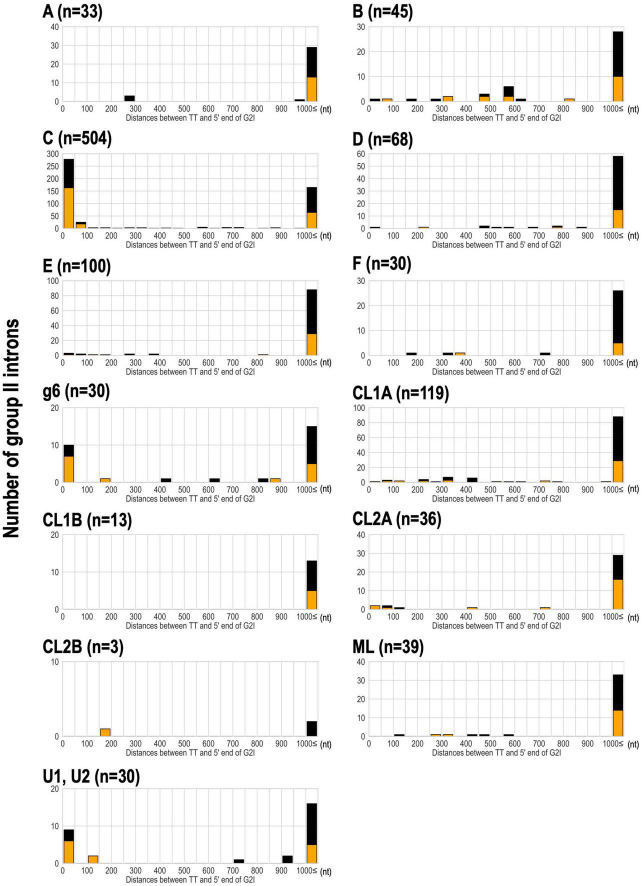
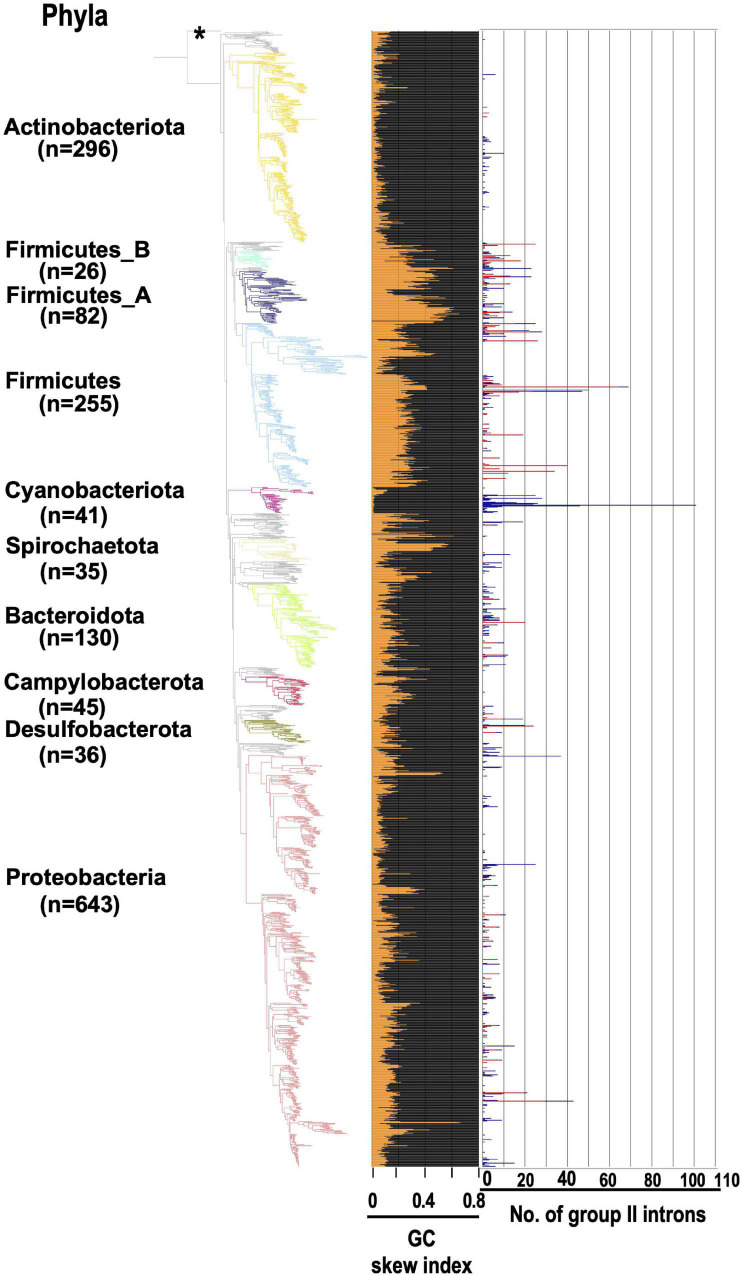
Group II introns (G2Is) are ribozymes that have retroelement characteristics in prokaryotes. Although G2Is are suggested to have been an important evolutionary factor in the prokaryote-to-eukaryote transition, comprehensive analyses of these introns among the tens of thousands of prokaryotic genomes currently available are still limited. Here, we developed a bioinformatic pipeline that systematically collects G2Is and applied it to prokaryotic genomes. We found that in bacteria, 25% (447 of 1,790) of the total representative genomes had an average of 5.3 G2Is, and in archaea, 9% (28 of 296) of the total representative genomes had an average of 3.0 G2Is. The greatest number of G2Is per genome was 101 in (phylum Cyanobacteriota). A comprehensive sequence analysis of the intron-encoded protein (IEP) in each G2I sequence was conducted and resulted in the addition of three new IEP classes (U1-U3) to the previous classification. This analysis suggested that about 30% of all IEPs are non-canonical IEPs. The number of G2Is per genome was defined almost at the phylum level, and at least in the following two phyla, Firmicutes, and Cyanobacteriota, the type of IEP was largely associated as a factor in the G2I increase, i.e., there was an explosive increase in G2Is with bacterial C-type IEPs, mainly in the phylum Firmicutes, and in G2Is with CL-type IEPs, mainly in the phylum Cyanobacteriota. We also systematically analyzed the relationship between genomic signatures and the mechanism of these increases in G2Is. This is the first study to systematically characterize G2Is in the prokaryotic phylogenies.
II组内含子(G2Is)是原核生物中具有反转录元件特征的核酶。尽管G2Is被认为是原核生物向真核生物转变过程中的一个重要进化因素,但目前对数万个原核生物基因组中这些内含子的全面分析仍然有限。在这里,我们开发了一种生物信息学流程,用于系统收集G2Is,并将其应用于原核生物基因组。我们发现,在细菌中,25%(1790个中有447个)的代表性基因组平均有5.3个G2Is,在古菌中,9%(296个中有28个)的代表性基因组平均有3.0个G2Is。每个基因组中G2Is数量最多的是蓝细菌门的一个物种,有101个。我们对每个G2I序列中的内含子编码蛋白(IEP)进行了全面的序列分析,结果在先前的分类基础上增加了三个新的IEP类别(U1 - U3)。该分析表明,所有IEP中约30%是非典型IEP。每个基因组中G2Is的数量几乎在门水平上就已确定,至少在以下两个门,即厚壁菌门和蓝细菌门中,IEP的类型在很大程度上是G2Is增加的一个相关因素,也就是说,具有细菌C型IEP的G2Is出现了爆发式增加,主要在厚壁菌门中,而具有CL型IEP的G2Is出现爆发式增加,主要在蓝细菌门中。我们还系统地分析了基因组特征与这些G2Is增加机制之间的关系。这是第一项在原核生物系统发育中系统表征G2Is的研究。