Institute for Research in Biomedicine (IRB Barcelona), The Barcelona Institute of Science and Technology, Barcelona, Spain.
Universitat Pompeu Fabra, Barcelona, Spain.
Elife. 2022 Apr 20;11:e75873. doi: 10.7554/eLife.75873.
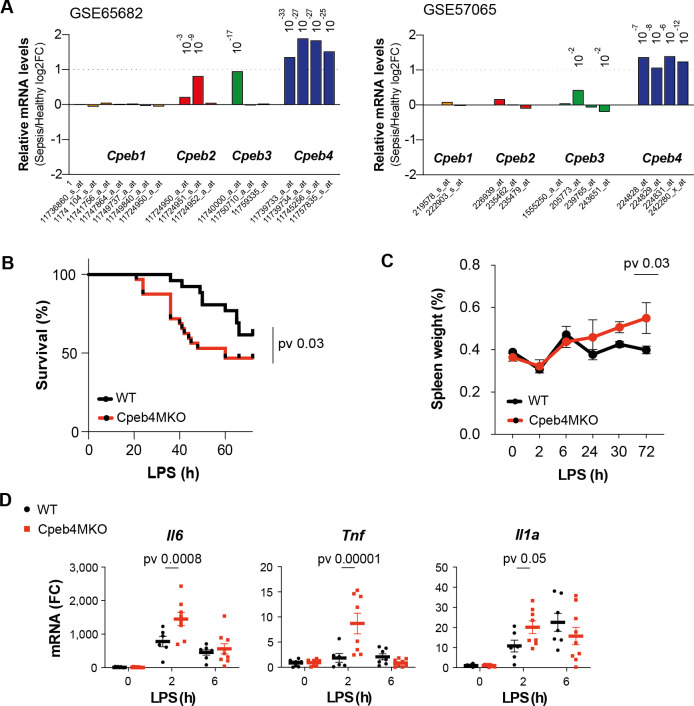
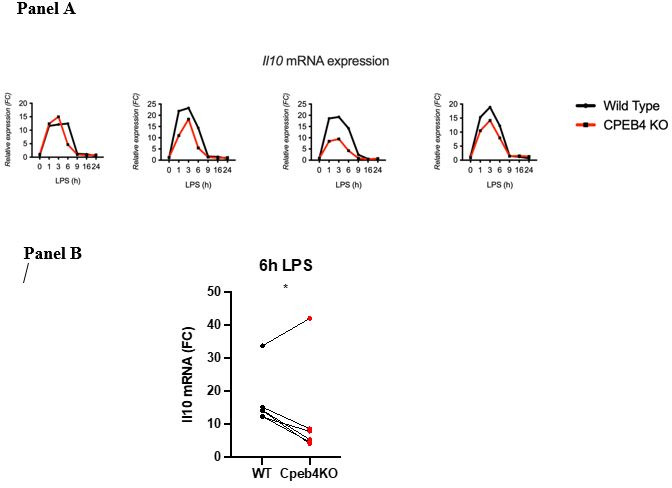
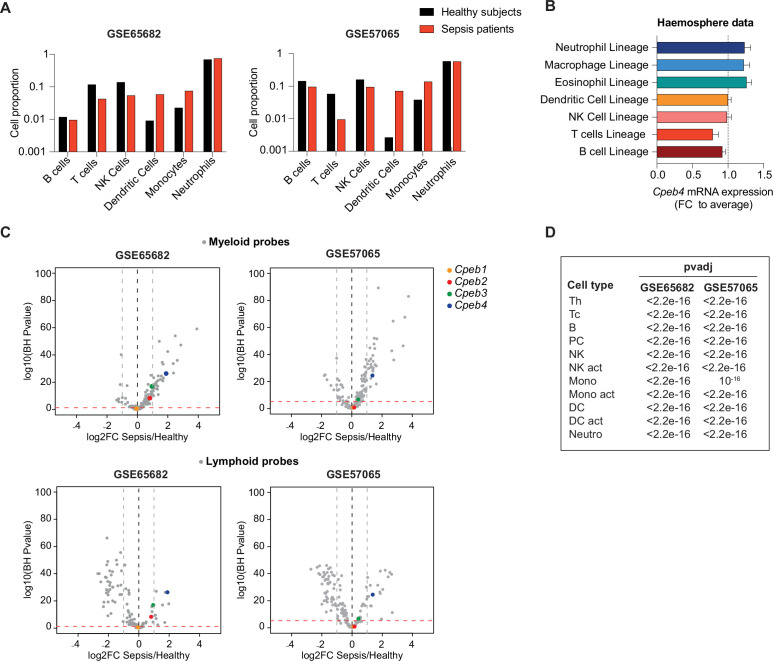
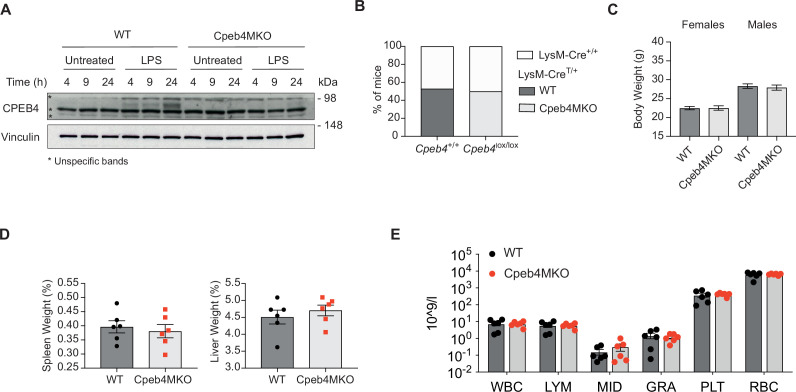
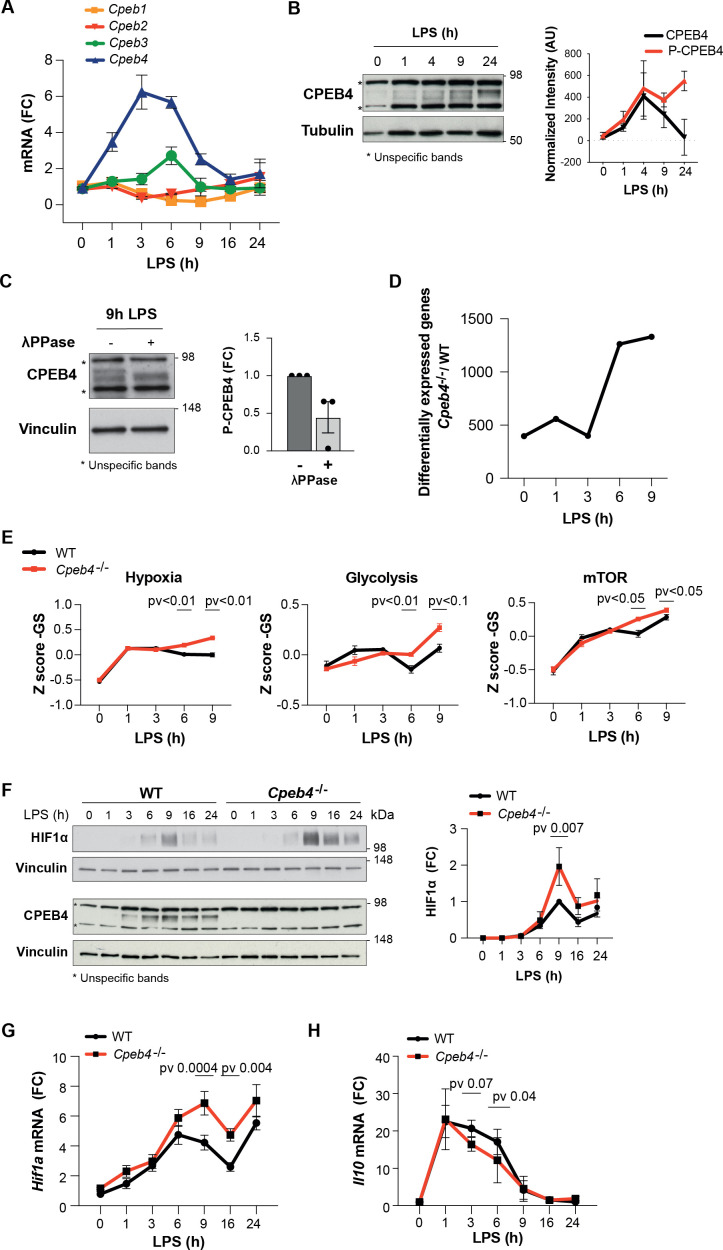
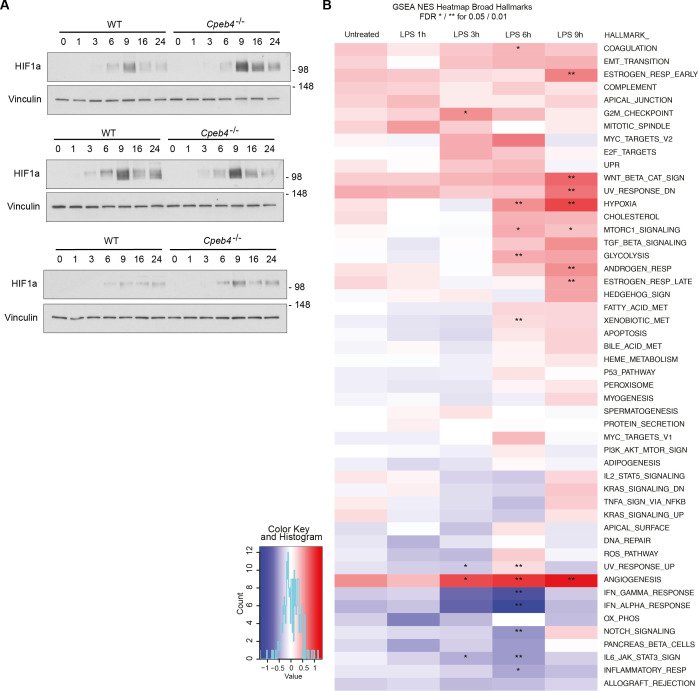
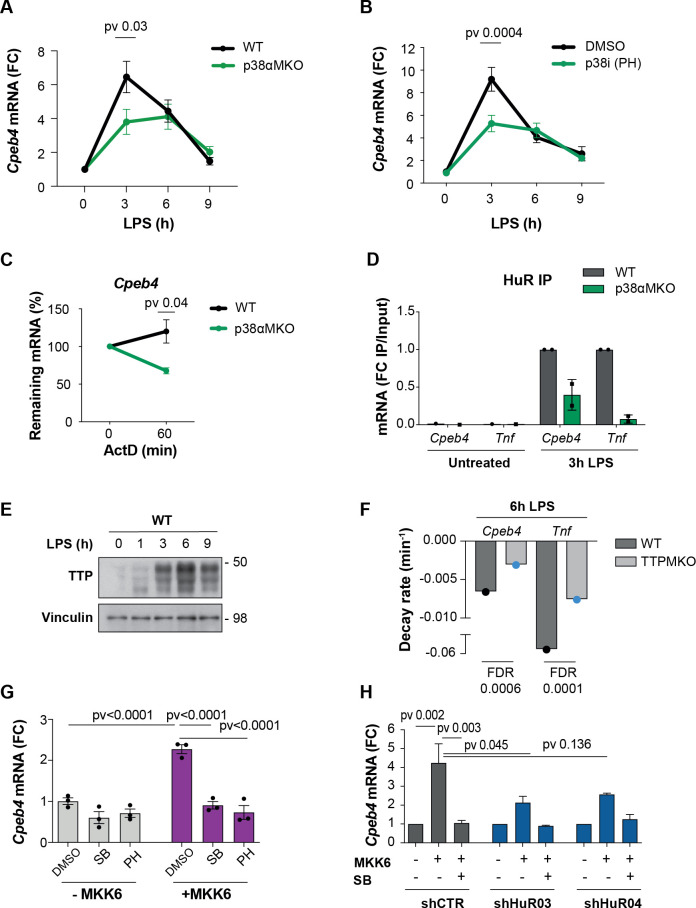
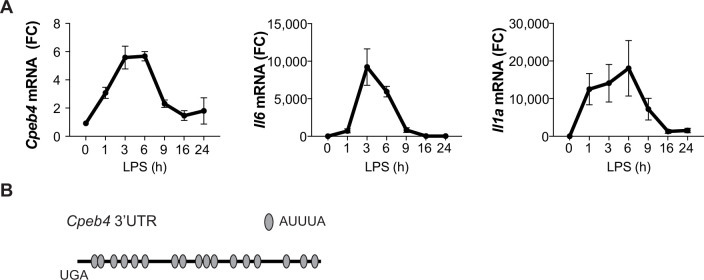
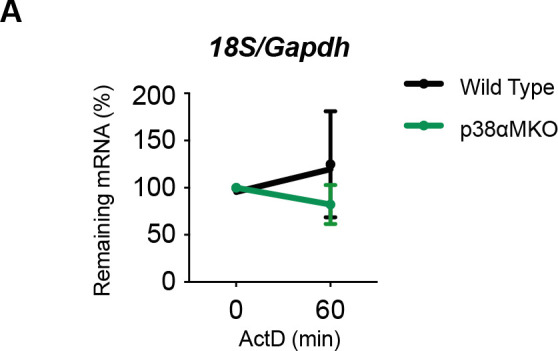
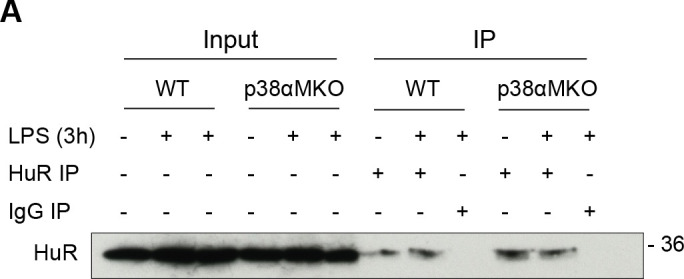
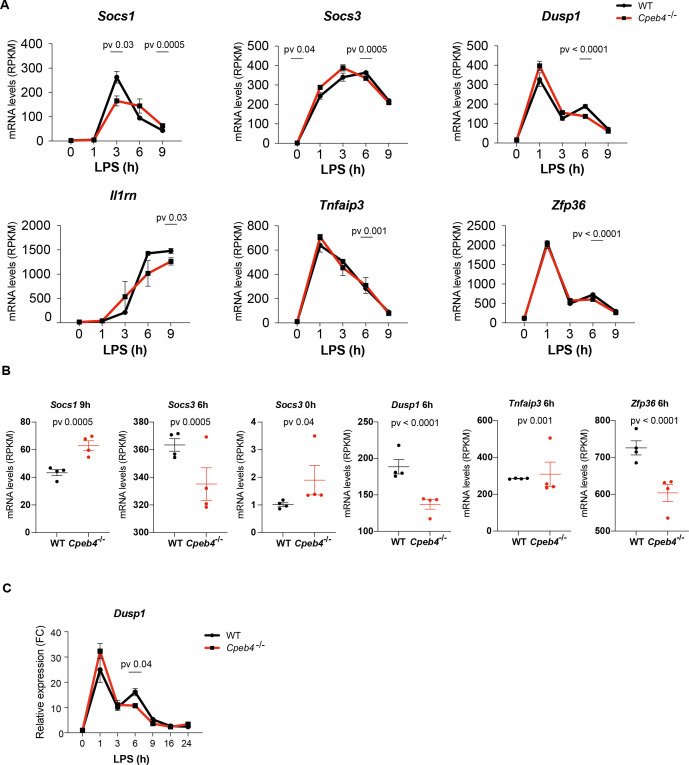
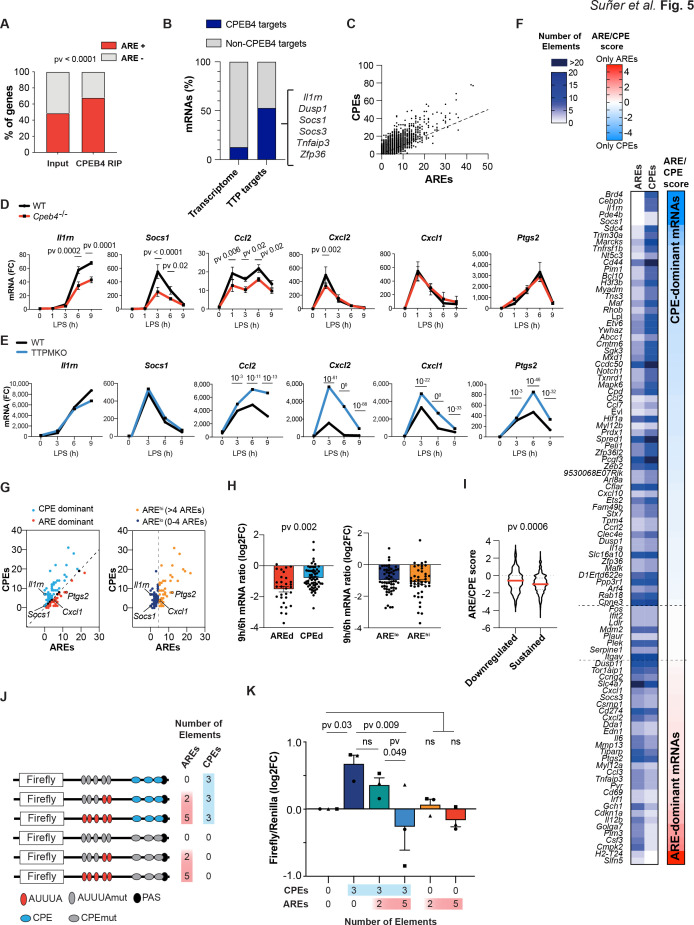
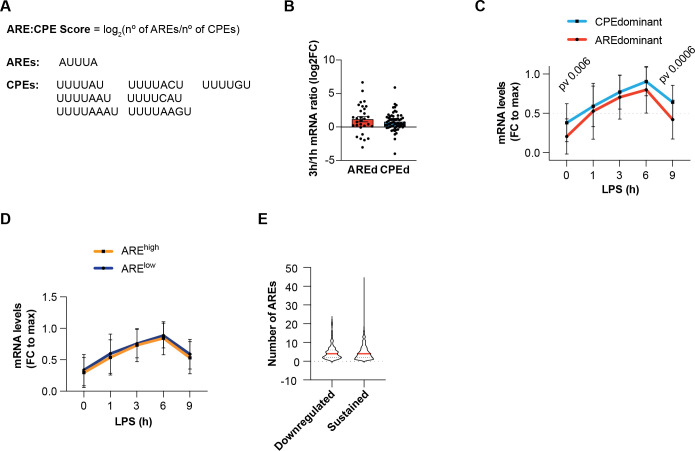
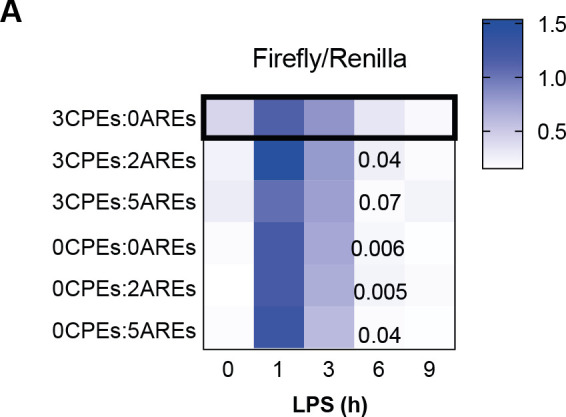
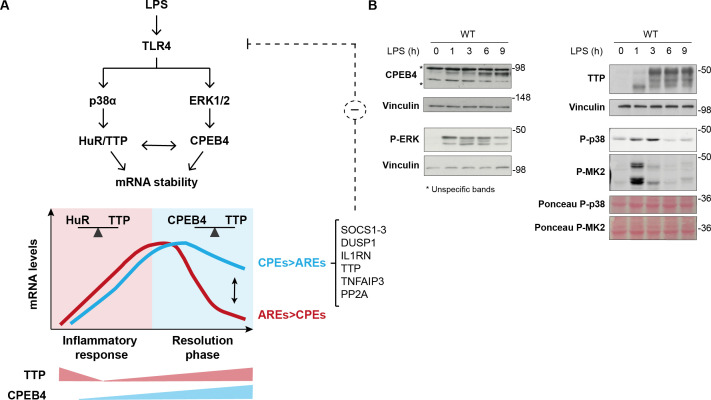
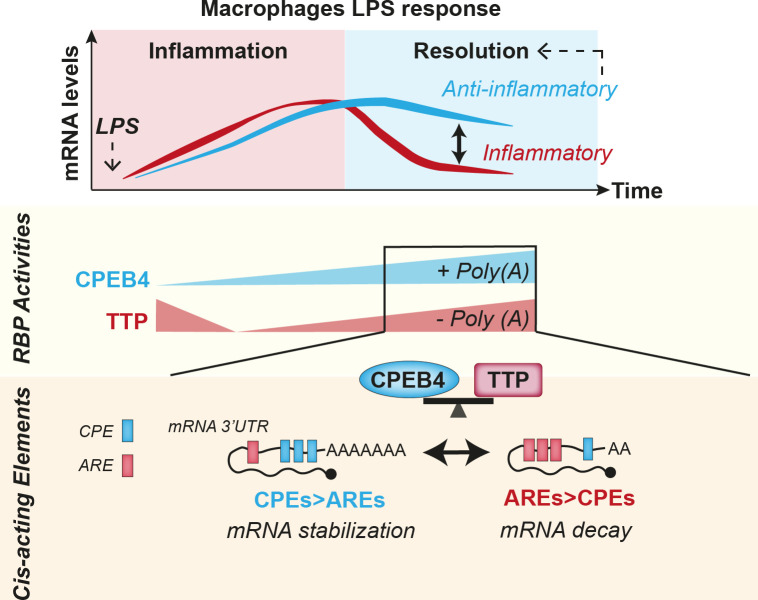
Chronic inflammation is a major cause of disease. Inflammation resolution is in part directed by the differential stability of mRNAs encoding pro-inflammatory and anti-inflammatory factors. In particular, tristetraprolin (TTP)-directed mRNA deadenylation destabilizes AU-rich element (ARE)-containing mRNAs. However, this mechanism alone cannot explain the variety of mRNA expression kinetics that are required to uncouple degradation of pro-inflammatory mRNAs from the sustained expression of anti-inflammatory mRNAs. Here, we show that the RNA-binding protein CPEB4 acts in an opposing manner to TTP in macrophages: it helps to stabilize anti-inflammatory transcripts harboring cytoplasmic polyadenylation elements (CPEs) and AREs in their 3'-UTRs, and it is required for the resolution of the lipopolysaccharide (LPS)-triggered inflammatory response. Coordination of CPEB4 and TTP activities is sequentially regulated through MAPK signaling. Accordingly, CPEB4 depletion in macrophages impairs inflammation resolution in an LPS-induced sepsis model. We propose that the counterbalancing actions of CPEB4 and TTP, as well as the distribution of CPEs and AREs in their target mRNAs, define transcript-specific decay patterns required for inflammation resolution. Thus, these two opposing mechanisms provide a fine-tuning control of inflammatory transcript destabilization while maintaining the expression of the negative feedback loops required for efficient inflammation resolution; disruption of this balance can lead to disease.
慢性炎症是疾病的主要原因。炎症反应的部分调节是通过编码促炎和抗炎因子的 mRNA 的不同稳定性来实现的。特别是,tristetraprolin(TTP)指导的 mRNA 去腺苷酸化使富含 AU 元件(ARE)的 mRNA 不稳定。然而,这种机制本身并不能解释为了使促炎 mRNA 的降解与抗炎 mRNA 的持续表达解偶联,所需的各种 mRNA 表达动力学。在这里,我们表明 RNA 结合蛋白 CPEB4 在巨噬细胞中与 TTP 以相反的方式发挥作用:它有助于稳定在其 3'-UTR 中含有细胞质多聚腺苷酸化元件(CPE)和 ARE 的抗炎转录本,并且它是脂多糖(LPS)触发的炎症反应消退所必需的。CPEB4 和 TTP 活性的协调通过 MAPK 信号通路进行顺序调节。因此,巨噬细胞中 CPEB4 的耗竭会损害 LPS 诱导的败血症模型中的炎症消退。我们提出,CPEB4 和 TTP 的拮抗作用,以及 CPE 和 ARE 在其靶 mRNA 中的分布,定义了炎症消退所需的转录特异性衰减模式。因此,这两种相反的机制提供了对炎症转录本不稳定的精细调节,同时维持了有效炎症消退所需的负反馈回路的表达;这种平衡的破坏可能导致疾病。