Center for Alzheimer's and Neurodegenerative Diseases, Peter O'Donnell Jr. Brain Institute, University of Texas Southwestern Medical Center, Dallas, Texas, USA.
Center for Alzheimer's and Neurodegenerative Diseases, Peter O'Donnell Jr. Brain Institute, University of Texas Southwestern Medical Center, Dallas, Texas, USA.
J Biol Chem. 2022 Jun;298(6):102014. doi: 10.1016/j.jbc.2022.102014. Epub 2022 May 5.
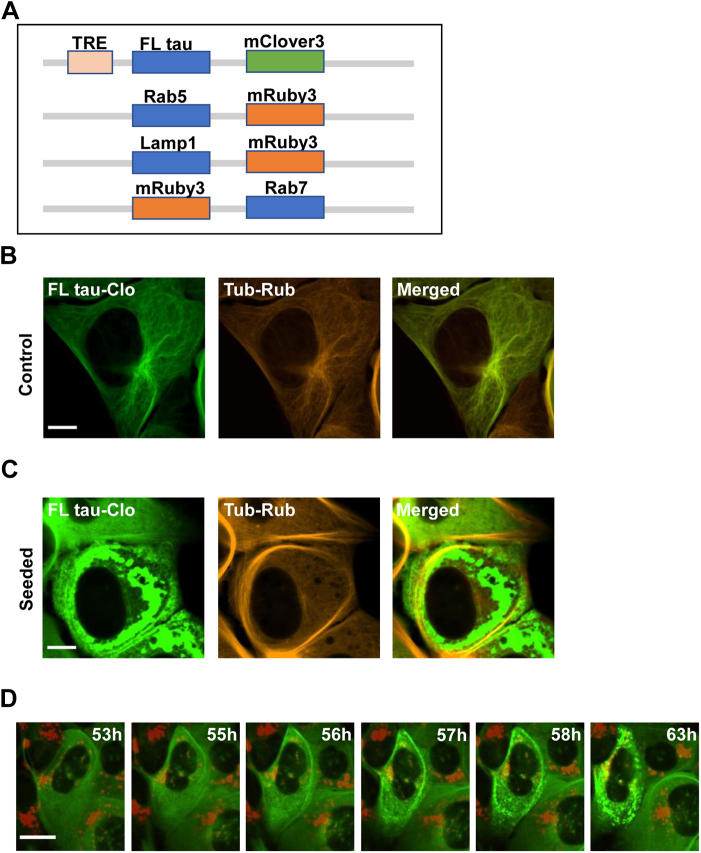
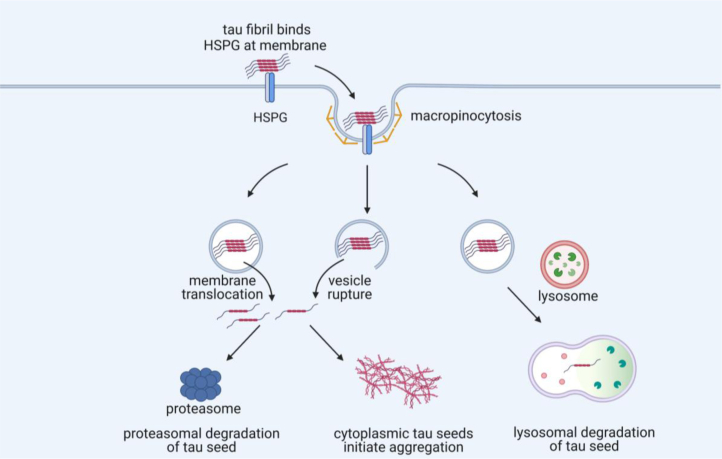
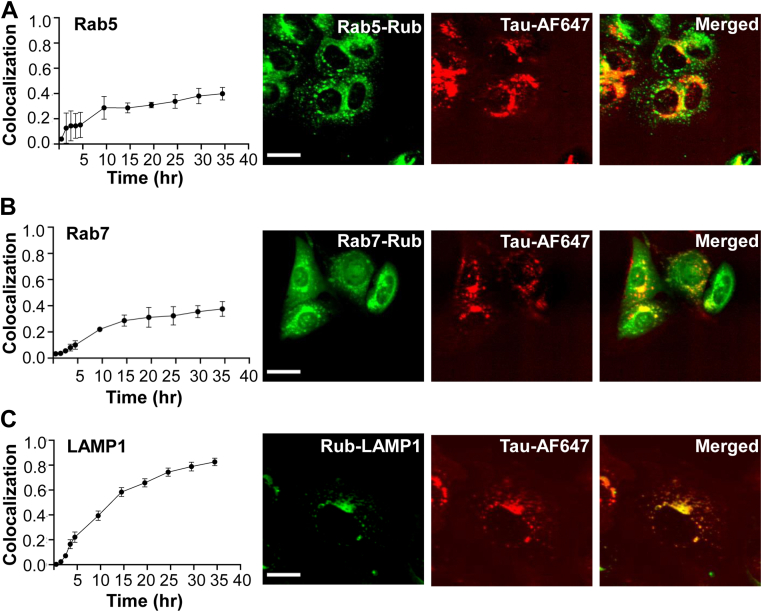
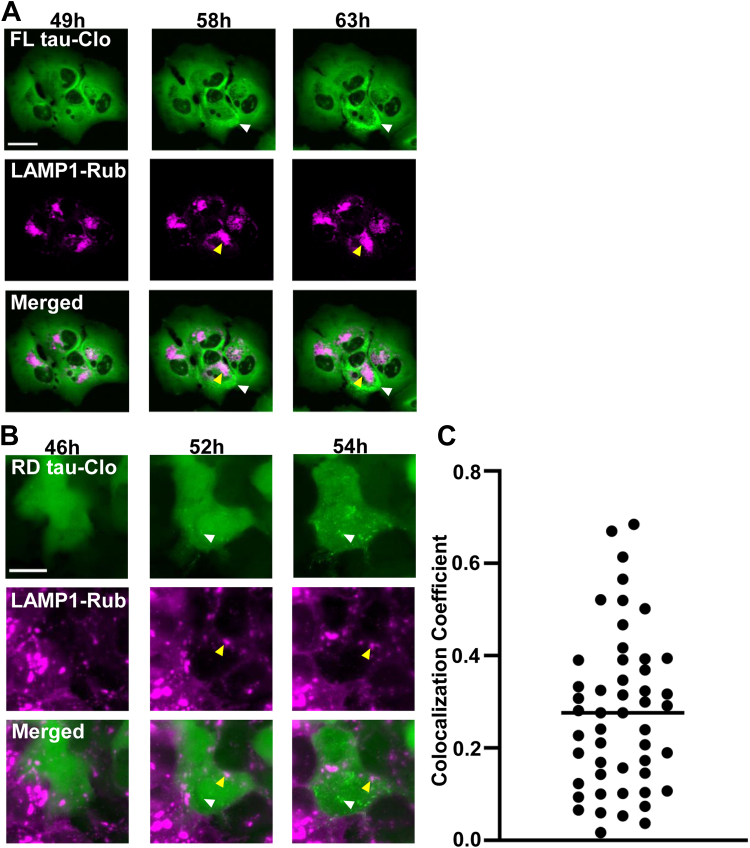
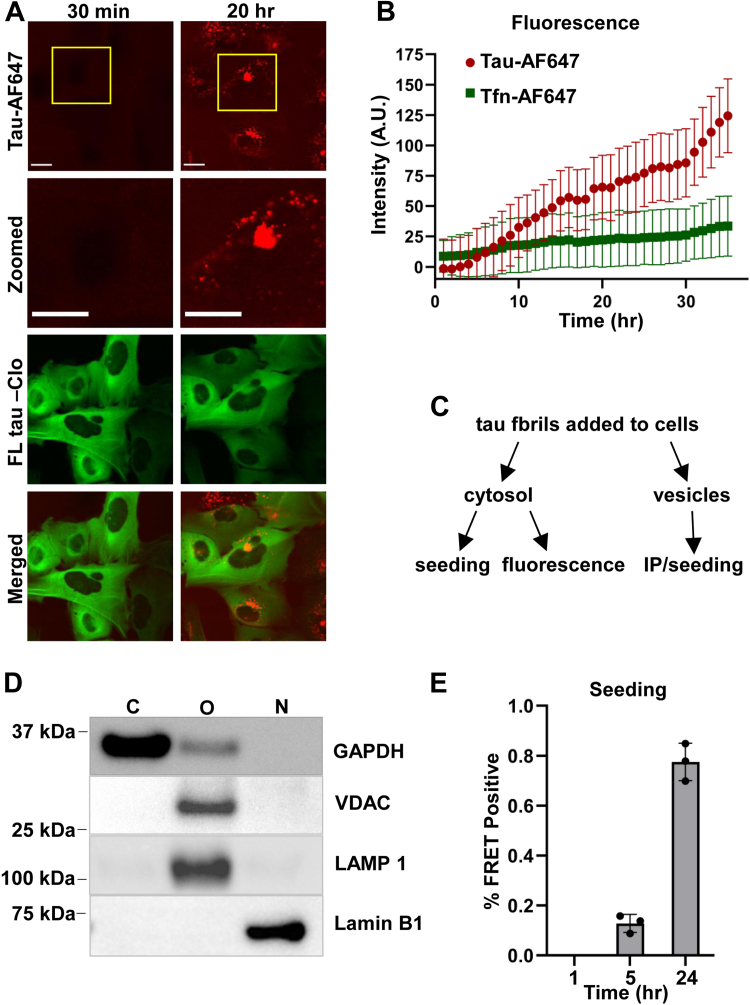
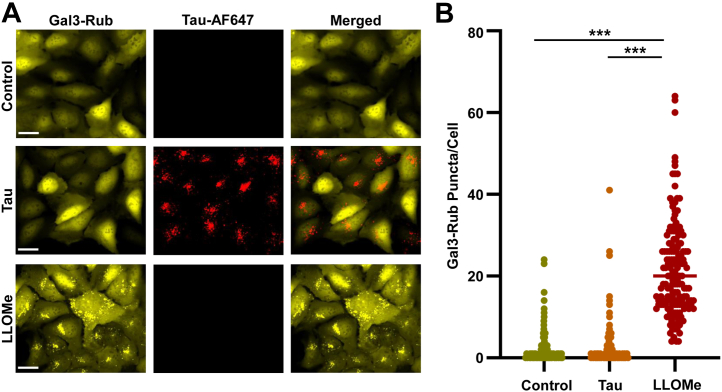
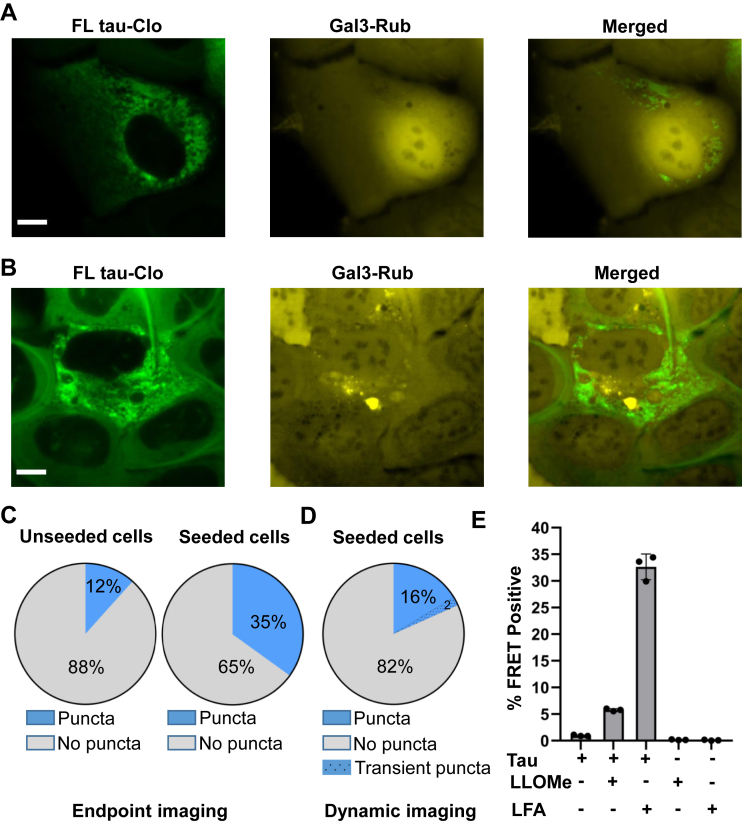
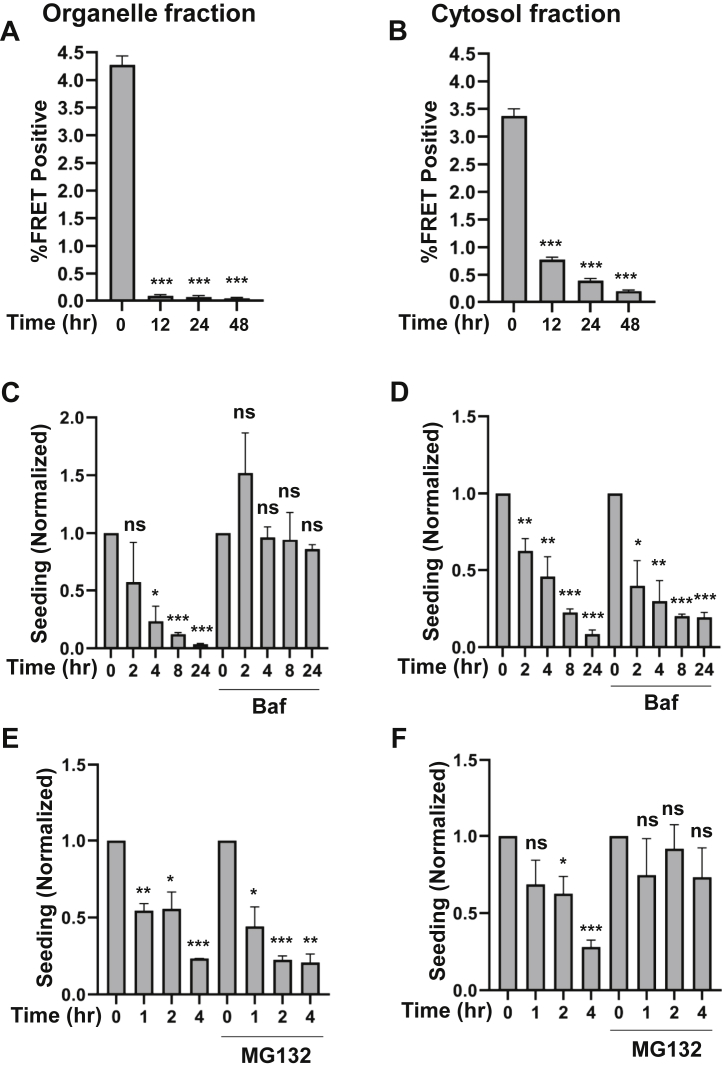
Tau assembly movement from the extracellular to intracellular space may underlie transcellular propagation of neurodegenerative tauopathies. This begins with tau binding to cell surface heparan sulfate proteoglycans, which triggers macropinocytosis. Pathological tau assemblies are proposed then to exit the vesicular compartment as "seeds" for replication in the cytoplasm. Tau uptake is highly efficient, but only ∼1 to 10% of cells that endocytose aggregates exhibit seeding. Consequently, we studied fluorescently tagged full-length (FL) tau fibrils added to native U2OS cells or "biosensor" cells expressing FL tau or repeat domain. FL tau fibrils bound tubulin. Seeds triggered its aggregation in multiple locations simultaneously in the cytoplasm, generally independent of visible exogenous aggregates. Most exogenous tau trafficked to the lysosome, but fluorescence imaging revealed a small percentage that steadily accumulated in the cytosol. Intracellular expression of Gal3-mRuby, which binds intravesicular galactosides and forms puncta upon vesicle rupture, revealed no evidence of vesicle damage following tau exposure, and most seeded cells had no evidence of endolysosome rupture. However, live-cell imaging indicated that cells with pre-existing Gal3-positive puncta were seeded at a slightly higher rate than the general population, suggesting a potential predisposing role for vesicle instability. Clearance of tau seeds occurred rapidly in both vesicular and cytosolic fractions. The lysosome/autophagy inhibitor bafilomycin inhibited vesicular clearance, whereas the proteasome inhibitor MG132 inhibited cytosolic clearance. Tau seeds that enter the cell thus have at least two fates: lysosomal clearance that degrades most tau, and entry into the cytosol, where seeds amplify, and are cleared by the proteasome.
tau 组装物从细胞外到细胞内空间的运动可能是神经退行性 tau 病的跨细胞传播的基础。这始于 tau 与细胞表面硫酸乙酰肝素蛋白聚糖结合,触发巨胞饮作用。然后,病理 tau 组装物被提出作为细胞质中复制的“种子”从囊泡隔室中逸出。tau 的摄取效率非常高,但只有约 1%到 10%的内吞聚集物的细胞表现出接种。因此,我们研究了添加到天然 U2OS 细胞或表达全长 (FL) tau 或重复结构域的“生物传感器”细胞中的荧光标记的全长 (FL) tau 原纤维。FL tau 原纤维与微管蛋白结合。种子在细胞质中同时在多个位置触发其聚集,通常与可见的外源性聚集物无关。大多数外源性 tau 转运到溶酶体,但荧光成像显示一小部分 tau 稳定积累在细胞质中。Gal3-mRuby 的细胞内表达,它与囊泡内的半乳糖结合,并在囊泡破裂时形成斑点,在 tau 暴露后没有显示出囊泡损伤的证据,并且大多数接种细胞没有内溶酶体破裂的证据。然而,活细胞成像表明,具有预先存在的 Gal3 阳性斑点的细胞的接种率略高于总体人群,这表明囊泡不稳定可能具有潜在的倾向性作用。tau 种子在囊泡和细胞质部分中的清除都很快发生。溶酶体/自噬抑制剂巴弗洛霉素抑制囊泡清除,而蛋白酶体抑制剂 MG132 抑制细胞质清除。因此,进入细胞的 tau 种子至少有两种命运:溶酶体清除,降解大多数 tau,以及进入细胞质,其中种子扩增,并被蛋白酶体清除。